
Manuscript accepted on :08-06-2026
Published online on: 19-06-2026
Plagiarism Check: Yes
Reviewed by: Dr. Saida Ncibi
Second Review by: Dr. Nataliya Kitsera and Dr. Priya Gayathri
Final Approval by: Dr. Hanefi ÖZBEK
Sardor Bakhtiyor ugli Sobirov1,3 , Abdisalim Abdikarimovich Zaripov1,5 , Inoyat Zulfiqorovich Zhumaev1* , Pulat Bekmuratovich Usmanov1 , Adilbay Tlepovich Esimbetov1 , Shavkat Yusubovich Rustamov1, Sadriddin Nurillo ugli Boboev1, Gulnaz Sultanbayevna Begdullaeva3, Eldor Bakhtiyor ugli Ibragimov1 , Shakhnoza Bakhtiyorovna Qurbonova1, Sadoqat Abdukhalil qizi Safarova2, Sherzod Niyatkobilovich Zhurakulov2and Dilbar Djamalovna Safarova4
, Abdisalim Abdikarimovich Zaripov1,5 , Inoyat Zulfiqorovich Zhumaev1* , Pulat Bekmuratovich Usmanov1 , Adilbay Tlepovich Esimbetov1 , Shavkat Yusubovich Rustamov1, Sadriddin Nurillo ugli Boboev1, Gulnaz Sultanbayevna Begdullaeva3, Eldor Bakhtiyor ugli Ibragimov1 , Shakhnoza Bakhtiyorovna Qurbonova1, Sadoqat Abdukhalil qizi Safarova2, Sherzod Niyatkobilovich Zhurakulov2and Dilbar Djamalovna Safarova4
1Department of Cell Biophysics, Institute of Biophysics and Biochemistry, National University of Uzbekistan, Tashkent, Uzbekistan.
2Department of Alkaloids Chemistry,Institute of the Chemistry of Plant Substances, Uzbek Academy of Sciences, Tashkent, Uzbekistan.
3Department of Animal Anatomy and Physiology, Karakalpak State University named after Berdakh, Nukus, Uzbekistan.
4Department of Medical and Biological Sciences, Uzbek State University of Physical Education and Sports, Chirchiq, Uzbekistan.
5Department of Physiology and Pathology, Tashkent State Medical University, Tashkent, Uzbekistan.
Corresponding Author E-mail: inoyat8585@mail.ru
DOI : https://dx.doi.org/10.13005/bpj/3476
Abstract
This study aims to evaluate how plasma membrane calcium transport mechanisms—specifically L-type calcium channels, receptor-operated calcium channels, and the sodium-calcium exchanger—contribute to the vasorelaxant activity of the alkaloid CAM-68. The isometric contraction force of the aortic rings was recorded using a Grass-Telefactor FT.03 myograph and analysed with the PowerLab data-acquisition software (ADInstruments). Findings indicate that the alkaloid CAM-68 exerts an effective vasorelaxant effect on vascular smooth muscle, which is primarily achieved by modulating intracellular Ca²⁺ concentration.The CAM-68 alkaloid inhibits voltage-dependent L-type Ca2+ channels and receptor-operated Ca2+ channels in the plasma membrane. It also attenuates the activity of the Na⁺/Ca²⁺ exchange in reverse mode. These effects are associated with the modulation of smooth muscle cell Ca²⁺ homeostasis and the attenuation of Ca²⁺ influx across the membrane.
Keywords
Alakaloid; L-type Ca2+-channels; Na⁺/Ca²⁺ exchange; Ouabain; Smooth muscle
Download this article as:| Copy the following to cite this article: Sobirov S. B. U, Zaripov A. A, Zhumaev I. Z, Usmanov P. B, Esimbetov A. T, Rustamov S. Y, Boboev S. N. U, Begdullaeva G. S, Ibragimov E. B. U, Qurbonova S. B, Safarova S. A. Q, Zhurakulov S. N, Safarova D. D. Evaluation of the Role of Plasmalemma Ca²⁺ Transport Systems in the Vasorelaxant Effect of the CAM-68 Alkaloid under In vitro Conditions. Biomed Pharmacol J 2026;19(2). |
| Copy the following to cite this URL: Sobirov S. B. U, Zaripov A. A, Zhumaev I. Z, Usmanov P. B, Esimbetov A. T, Rustamov S. Y, Boboev S. N. U, Begdullaeva G. S, Ibragimov E. B. U, Qurbonova S. B, Safarova S. A. Q, Zhurakulov S. N, Safarova D. D. Evaluation of the Role of Plasmalemma Ca²⁺ Transport Systems in the Vasorelaxant Effect of the CAM-68 Alkaloid under In vitro Conditions. Biomed Pharmacol J 2026;19(2). Available from: https://bit.ly/4guZlV2 |
Introduction
Arterial hypertension serves as a primary driver of cardiovascular disease and remains a leading risk factor for both myocardial infarction and stroke.1Therefore, based on the latest advances in molecular pharmacology, developing novel approaches to the adequate treatment of arterial hypertension is one of the pressing issues in modern.2,3Information on the pharmacological properties of key vascular targets directly involved in the pathogenesis of arterial hypertension is considered of significant importance.In this case, the primary focus is on elucidating the mechanisms of pharmacological modulation of Ca2+ transport systems in smooth muscles. Furthermore, the intracellular Ca²⁺ concentration performs a significant function in the neurohumoral governance of vascular tension.4Within vascular smooth muscle cells (VSMCs), the processes of contraction and relaxation are primarily determined by changes in intracellular Ca²⁺ ion concentration ([Ca²⁺]i).5-7Therefore, the influx and efflux of Ca²⁺ ions through the plasma membrane play a key role in the regulation of blood vessel tone.8,9Among alkaloids, diterpenoid alkaloids are considered to have the highest antiarrhythmic activity. In particular, it has been noted that lappakonitine has strong antiarrhythmic properties.Numerous studies have shown that diterpenoid alkaloids possess vasorelaxant properties, and their mechanism of action is thought to be closely linked to membrane systems that regulate Ca²⁺ homeostasis.10-12In particular, reducing the activity of voltage-dependent L-type Ca²⁺-channelsand enhancing Ca²⁺ extrusion via the Na⁺/Ca²⁺ exchanger (NCX) are regarded as potential mechanisms of action of the alkaloids.13-15
Therefore, the aim of this study is to determine the role of plasma membrane Ca²⁺ transport systemsnamely Ca²⁺L-channels, Ca²⁺R-channels and the NCXin the vasorelaxant effect of the alkaloid 1-Ethyl-7α,11α-dihydroxy-6,8,10-trimethoxydodecahydro-2H-3,6a, 12-(epietane[1,1,2]triyl)-7,9-methanoneftho[2,3-b]azocin-3(4H)-yl-2-(ethylsulphonamido) benzoate (CAM-68). A thorough analysis of these mechanisms will provide the scientific basis for developing new natural vasodilator agents and innovative approaches to the treatment of vascular diseases.16
The isolation of CAM-68
Continuing our work on the modification of the anthranilic acid moiety of lappakonitine (1), we obtained N-deacetyl-lappaconitine (2) (83%) using a modified method.17Reaction 2 with ethylsulphonyl chloride was carried out using three methods,18 yielding the corresponding ethylsulphonamide derivatives 3 in 86% yield.
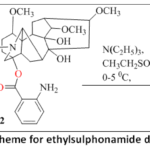 |
Scheme 1: Synthesis scheme for ethylsulphonamide derivatives of 3-lappaconitine. |
The structures of the synthesised compounds 2 and 3 were confirmed using 1H and 13C NMR spectroscopy.
Product 3’s 1H NMR spectrum shows signals in the high-field region that are indicative of two methyl groups: δ 1.10 (NCH₂Me) and δ 1.33 (SO₂CH₂Me). In the δ 3.28–3.40 m, singlets corresponding to the protons of methoxyl (-OCH₃) groups linked to carbons C-1, C-14, and C-16 were found.d. area. A singlet signal was detected at δ 10.48 in the aromatic portion of the spectrum, which corresponds to an NH group.
The carbonyl group carbons, the benzene ring carbons, and the three quaternary carbons bound to oxygen are represented by signals in the 13C NMR and HSQC spectra. A signal was detected at δ 167.30 ppm for the carbonyl group carbon (C=O), δ 141.18 ppm for the C-2′ carbon, and δ 116.29 ppm for the C-1′ carbon in the low-field area of the spectrum. The higher-frequency regions showed signals related to carbon atoms bound to oxygen: δ 58.03 (14-OCH₃), δ 56.63 (1-OCH₃), and δ 56.24 (1-OCH₃). The carbon atoms 13C NMR signals show no additional modifications as compared to 2. These spectrum data were used to fully identify the structure of the molecule 1-ethyl-7a,11a-dihydroxy-6,8, 10-trimethoxydodecahydro-2H-3,6a,12-(epant[1,1,2]triyl)-7,9-methanonaphth[2,3-b]azocyclo-3(4H)-yl-2-(ethylsulphonamidobenzoate) (3).
An FT-IR/NIT Spectrum 3 spectrometer equipped with a frustrated total internal reflection (FTIR) system was used to record the infrared spectra. A JNM-ECZ600R spectrometer (JEOL, Japan) was used to record the NMR spectra for 1H in CDCl3 solutions at an operating frequency of 600 MHz. The internal standard in the ¹H NMR spectra was TMS (0 m.d.). The internal standard in the 13C NMR spectra at an operating frequency of 150 MHz was the solvent’s chemical shift (CDCl₃, 77.16 ppm relative to TMS). An Agilent Technologies 6420 mass spectrometer equipped with a triple quadruple LS/MS system (electrospray ionization, ESI TIC Scan, and CAMAG TLC-MS with an ACQUITY QDa detector) was used to record the HR-ESI-MS data. Using a variety of solvent solutions, TLC on Sigma-Aldrich plates (Silufol L/W, 10 cm × 20 cm, with a 254 nm fluorescent indicator, Germany) was used to track the reaction’s progress and the purity of the chemicals produced. A BOETIUS micro-stolich was used to determine the melting points of all the synthesized materials.
Ethyl 7α,11α-dihydroxy-6,8,10-trimethoxydodecahydro-2H-3,6α,12-(epant[1,1,2]triyl)-7,9-methanonaphth[2,3-b]azocyclo-3(4H)-yl 2-aminobenzoate (2).
0.884 ml of 1 N HCl solution (30.7 mmol) was added to 2.0 g (3.4 mmol) of lappaconitine (1) in 9.4 ml of distilled water. For two hours, the reaction was conducted in a water bath between 95 and 100 °C. TLC was used to track the reaction’s development. N-deacetyl-lappaconitine was extracted using chloroform, dried over Na₂SO₄, and then concentrated by evaporation after the pH was adjusted to 8 using a saturated solution of NaHCO₃. After the precipitate was recrystallized from methanol, 1.545 g (83%) of colorless crystals with a melting point of 215–217 °C (methanol) and Rf 0.45 (benzene:chloroform:triethylamine 4:1:0.3) were obtained.
1-Ethyl-7α,11α-dihydroxy-6,8, 10-trimethoxydodecahydro-2H-3,6a,12-(epietane[1,1,2]triyl)-7,9-methanoneftho[2,3-b]azocin-3(4H)-yl-2-(ethylsulphonamido)benzoate (3).
2.0 g (3.68 mmol) N- desacetyllappaconitin (2) was dissolved in 20 ml of dichloromethane and, whilst stirring at 0–5 °C, 1.43 g (14.1 mmol) of triethylamine and 0.5 ml (5.28 mmol) of ethyl sulphonyl chloride were added gradually. The mixture was stirred for 1 hour at a temperature of 0–5 °C. The reaction mixture was then stirred at room temperature for 1 hour (TLC control). The reaction product was washed twice with an aqueous solution of NaHCO₃, and then with a 20% aqueous solution of NaCl. The CH₂Cl₂ solution was dried over Na₂SO₄; the resulting residue was evaporated to dryness and recrystallised from methanol, yielding 2.0 g (86%) of product 3. C32H46N2O9S – colourless crystals, melting point 190–192 °C (methanol), Rf 0.69 (benzene: chloroform: triethylamine, 4:1:0.3).
1H NMR (600 MHz,CDCl3, δ, ppm, J/Hz): 10.48 (1H, s, SO2NH), 7.92 (1H, dd, J = 8.0, 1.6, H-6′), 7.70 (1H, dd, J = 8.4, 1.1, H-3′), 7.48 (1H, ddd,J = 8.6, 7.3, 1.7, H-4′), 7.04 (1H, ddd, J = 8.2, 7.3, 1.1, H-5′), 3.58 (1H, d, J = 11.4, Ha-19), 3.52 (1H, s, 9-OH), 3.43 (1H, dd,J = 4.9, 1.1, Hb-19), 3.40 (3H, s, 14-OCH3), 3.30 (4H, br.s, 16-OCH3, SO2CH2Me), 3.28 (3H, s, 1-OCH3), 3.20–3.11 (3H, m, H-1,7, 10), 2.99 (1H, s, 8-OH), 2.70–2.63 (2H, m, CHaCH3, H-16), 2.59–2.45 (4H, m, NCHbCH3, Ha-6, H-14, 17), 2.41–2.35(3H, m, Ha-3, H-5, 13), 2.30–2.23 (2H, m, Hb-3, Ha-15), 2.19–2.14 (2H, m, Hb-15, Ha-12), 2.08 (1H, dd, J = 12.5, 4.7, Hb-12),2.02–1.94 (2H, m, Hb-6, Ha-2), 1.77 (1H, dddd, J = 14.5, 12.1, 4.9, 2.3, Hb-2), 1.57–1.54 (1H, m, SO2CH2Me), 1.33 (3H, t,J = 7.4, SO2CH2Me), 1.10 (3H, t, J = 7.1, NCH2Me).
13C (150 MHz, CDCl3, δ, ppm): 167.30 (OCO), 141.18 (C-2′), 134.67 (C-4′),131.80 (C-6′), 122.59 (C-3′), 117.90 (C-5′), 116.29 (C-1′), 90.23 (C-14), 85.25 (C-4), 84.18 (C-1), 82.97 (C-16), 78.66 (C-9),75.76 (C-8), 61.61 (C-17), 58.03 (14-OCH3), 56.63 (1-OCH3), 56.24 (16-OCH3), 55.47 (C-19), 51.08 (C-11), 49.90 (NCH2CH3), 49.08 (C-5), 48.69 (C-7), 47.64 (SO2CH2CH3), 46.55 (C-13), 45.00 (C-15), 36.38 (C-10), 31.87 (C-3), 26.87 (C-6), 26.29 (C-2), 24.23 (C-12), 13.62 (NCH2CH3), 8.23 (SO2CH2CH3).
ESI-MS m/z 635.6562 [M+Н]+ (calcd for C32H47N2O9S).
Materials and Methods
The trials were conducted on aortic preparations from white, inbred rats (200–250 grams). The animals were anaesthetised with sodium pentobarbital, their thoracic cavities were opened, and the aorta was surgically isolated. The surrounding connective tissue and adipose layer were carefully removed, and the aorta was divided into 3–4 mm-long cylindrical rings.19The rings were placed in a specialised 5 ml organ bath and perfused with Krebs–Henseleit physiological solution (mM): NaCl 120.4; KCl 5.0; NaHCO₃ 15.5; NaH₂PO₄ 1.2; MgCl₂ 1.2; CaCl₂ 2.5; C₆H₁₂O₆ 11.5; HEPES buffer, pH 7.4). The warmth within the organ bath was kept at 37 °C utilizing an LK LAB (Korea) ultrathermostat, and the solutions were continuously aerated with carbogen (95% O₂, 5% CO₂).20Each aortic ring was connected to a Grass FT.03 (Grass-Telefactor, USA) sensor via two stainless-steel platinum hooks, the signal was transmitted to an amplifier and recorded using PowerLab (ADInstruments) software for analysis on a computer.Initially, each ring was subjected to an isometric force of 1 g (~10 mN) and equilibrated for 60 minutes.After the equilibration period, experiments were conducted to assess the vasorelaxant properties of the alkaloid CAM-68.
All chemical agents (verapamil, phenylephrine, ouabain) were of analytical quality and were procured from Sigma Ltd Co. (St. Louis, MO, USA).The alkaloid CAM-68 was provided by researchers from the Institute of Plant Chemistry of the Uzbek Academy of Sciences. All investigations were carried out in complete adherence to international ethical standards and rules concerning the employment of laboratory creatures.
Experimental Design
Aortic ring contraction responses were elicited using potassium chloride (KCl) or the α1-adrenoceptor agonist phenylephrine (PE). The maximum contraction produced by KCl or PE was taken as 100% as the control.21Once the resulting contractions had reached their maximum, the dose-dependent relaxing effect of the alkaloid CAM-68 was examined.
Assessment of the involvement of L-type Ca²⁺ channels.To assess the contribution of L-type Ca2+ channels to the vasorelaxant effect of the alkaloid CAM-68, the preparation was incubated with the channel-selective blocker verapamil prior to contraction induction. Once the contraction had stabilised, the relaxant effect of CAM-68 was examined and compared with the responses obtained in the absence of the blocker. Simultaneously, the function of Ca²⁺ channels was further established in further trials by adjusting the CaCl₂ level (0.5–2.5 mM) within the Krebs–Henseleit fluid.The function of voltage-gated Ca²⁺ channels in the mechanism of action of this alkaloid was evaluated by modifying the extracellular Ca²⁺ concentration.
Assessment of the involvement of receptor-operated Ca2+-сhannels (ROCCs).To assess the receptor-mediated modulation of Ca2+ channels by the alkaloid CAM-68, the preparation was incubated with phentolamine before PE-induced contraction. After the contraction force reached its maximum, the relaxing effect of the CAM-68 was compared with that of the control.
Evaluation of the participation of NCX.The effect of the alkaloid CAM-68 on NCX activity was assessed in Krebs solution without Na⁺ ions, relative to slowly occurring contractions. The dose-dependent relaxing effect of alkaloid CAM-68 on contractions induced by the reverse-mode influx of Ca2+ ions into the cell was studied. Similarly, the effect of the alkaloid CAM-68 on NCX was achieved by modulating the Na⁺/K⁺-ATPase. That is, the relaxing effect on slowly developing contractions with the aid of ouabain was examined.
Statistical analysis
The contraction responses of blood vessel smooth muscles were calculated as a percentage (%) of the maximum contraction value. The IC50value was determined relative to the 50% reduction in maximal contraction on the concentration-response curve for the alkaloid. The study findings were derived from n- separate trials and are shown as M±m, where M- is the average value and m- is the standard deviation. The information was evaluated for typical distribution using the Shapiro–Wilk test and for variance consistency using Levene’s test. Statistical importance was determined at p<0.05 and p<0.01. All computations were executed utilizing the OriginPro 2021 (OriginLab Corporation, USA) analytical program.
Results
The effect of the alkaloid CAM-68 on L-type Ca2+-channels
In initial trials, it was observed that the alkaloid СAM-68 exhibits a dose-dependent vasorelaxant action on KCl-induced aortic ring contractions.It was found that the alkaloid СAM-68 at a concentration of 5 μM reduced 50 mM KCl-induced aortic contraction by 14.8±3.7%, and at 50 μM by 92.1±3.8%. Under these conditions, the IC₅₀ value of alkaloid СAM-68 was 17.9 μM(Figure 1).
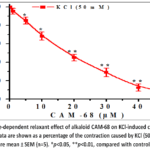 |
Figure 1: Dose-dependent relaxant effect of alkaloid СAM-68 on KCl-induced contraction of aortic rings. |
The results obtained showed that the СAM-68 alkaloid has a relaxing effect on KCl-induced aortic contractions. From this, it can be hypothesized that the vasorelaxant impact of the СAM-68 alkaloid is linked to blocking the L-type Ca²⁺-channel and a lowering of the [Ca²⁺]i level.In subsequent experiments, the vasorelaxant effect of CAM-68 in Krebs–Henseleit solution was analysed by varying the CaCl₂ concentration, i.e. with [Ca²⁺]out in the range of 0.1–2.5 mM. Under these conditions, pretreatment with the alkaloid СAM-68 was found to reduce the CaCl₂ concentration-dependent contractile force of aortic rings (Figure 2, A). That is, incubation with alkaloid СAM-68 (50 μM) reduced the force of contraction elicited by the maximal concentration of CaCl₂ in aortic rings by 79.7±3.8 % compared with control. The present effect of the alkaloid CAM-68 likely indicates inhibition of voltage-dependent L-type Ca²⁺-channelslocated in the plasma membrane. To identify this mechanism, the effect of the alkaloid CAM-68 was compared with that of verapamil, a selective blocker of L-type Ca²⁺-channels.In experiments, verapamil (1 µM) was found to reduce the CaCl₂-dependent contractile force of aortic rings by 92.6±3.9% compared with the control group. These results confirmed that verapamil is an effective blocker of voltage-dependent L-type Ca²⁺-channels. At the same time, the inhibitory effect of the alkaloid CAM-68 was observed to be less pronounced than that of verapamil. The combined effect of the alkaloid CAM-68 with verapamil was also studied. In experiments, incubation with verapamil (IC₅₀=0.1 μM) was observed to reduce the KCl-induced contractile force of aortic rings by 50±3.9% compared with control. Under these conditions, addition of the alkaloid CAM-68 (IC₅₀=17.9 μM) was found to reduce the contraction force by a further 27.5±4.1% (Figure 2B).
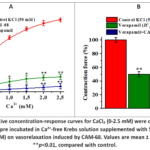 |
Figure 2: A-Cumulative concentration-response curves for CaCl₂ (0-2.5 mM) were constructed in aortic rings that has been pre incubated in Ca²⁺-free Krebs solution supplemented with 50 mM KCl. B- effect of verapamil (0.1 μM) on vasorelaxation induced by CAM-68. |
The results obtained showed that the alkaloid CAM-68 attenuates Ca²⁺ entry into the cell by inhibiting the activity of voltage-dependent L-typeCa²⁺-channels. However, under conditions of blockade by verapamil, the additional vasorelaxant effect of the alkaloid CAM-68 may be associated with other ion transport systems.
The effect of the CAM-68 alkaloid on ROCCs
The contraction that occurs under PE-induced conditions is supplied by Ca2+ ions released from the sarcoplasmic reticulum and entering via receptor-operated Ca2+ channels (ROCCs).22In experiments, it was found that the contraction force of the aortic ring induced by PE was reduced by 9.5±3.4% compared with control at 10 μM alkaloid CAM-68, and by 89.8±3.8% at 150 μM. Under these conditions, the IC₅₀ value of the CAM-68 alkaloid was 55.4 μM. In later trials, we performed research with the α-adrenergic receptor antagonist phentolamine to examine the impact of the alkaloid on ROCCs. During phentolamine incubation, the vasorelaxant action of the alkaloid CAM-68 on the contractile strength of an aortic sample provoked by PE (1 μM) was diminished by 34.3±3.9% relative to control. (Figure 3).
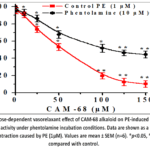 |
Figure 3: Dose-dependent vasorelaxant effect of СAM-68 alkaloid on PE-induced aortic ring contraction activity under phentolamine incubation conditions. |
The experiments conducted suggest that the vasorelaxant effect of the alkaloid СAM-68 is associated with the blockade of ROCCs, a conclusion confirmed by the results of experiments carried out in the presence of the α-adrenoceptor blocker phentolamine.
Effect of the alkaloid CAM-68 on NCX
In the next experiments, the effect of the CAM-68 alkaloid on NCX was studied. In aortic rings in Krebs solution without Na⁺ ions, slowly developing contractions were observed.It is known that a reduction in the external Na⁺ concentration promotes the reverse mode of the NCX, aiding the influx of Ca²⁺ ions into the cell, and this results in the beginning of contraction.23Under these conditions, alkaloid the CAM-68 (150 µM) significantly reduced the contractile force in aortic rings, with a decrease of 61.6±3.9 % compared with the control group. We also observed the effects of these alkaloids in the presence of KB-R7943 (5 μM), a specific NCX blocker. In experiments, under KB-R7943 incubation conditions, the contraction of Krebs solution devoid of Na⁺ ions was reduced from 61.6±3.9% to 32.7±3.7% by the alkaloid CAM-68 (Figure 4A). The findings gathered suggest that the vasorelaxant action of the CAM-68 alkaloid is linked to inhibition of NCX activity in reverse mode. Additionally, to further clarify the effect of the alkaloid CAM-68 on NCX, experiments were conducted in the presence of the Na⁺/K⁺-ATPase inhibitor ouabain. Under the influence of ouabain (10 μM), the Na⁺/K⁺ -ATPase activity is reduced, resulting in an increase in intracellular Na⁺ concentration, which in turn leads to Ca²⁺ entry into the cell and enhanced contraction due to the reverse mode of operation of the NCX.In this study, the alkaloid CAM-68 (150 µM) was found to reduce the ouabain-induced contraction by 68.7±4.2% compared with control. Under these conditions, incubation with KB-R7943 (5 μM) attenuated the effect of the alkaloid CAM-68 from 68.7±4.2% to 36.4±4.0% (Figure 4B). Overall, the results indicate that the vasorelaxant effect of the alkaloid CAM-68 is achieved by inhibiting the reverse-mode activity of NCX.
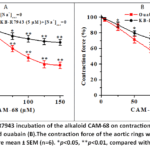 |
Figure 4: Effect of KB-R7943 incubation of the alkaloid CAM-68 on contractions induced by Na⁺-free Krebs solution (A) and ouabain (B).The contraction force of the aortic rings was taken to be 100%. |
The alkaloid CAM-68 can reduce the concentration of Ca2+ ions in smooth muscle cells by inhibiting NCX activity, thereby promoting cell relaxation.
Discussion
Diterpenoid alkaloids, as natural compounds that modulate cellular Ca²⁺ homeostasis, play an important role in controlling vasomotor functions in vascular smooth muscle by acting on ion channels, ion transport systems and signalling pathways.24Among these compounds, several diterpenoid alkaloids have been reported to exert vasorelaxant effects through the modulation of multiple Ca²⁺-dependent mechanisms, including voltage-dependent Ca²⁺ channels, receptor-operated pathways, and intracellular Ca²⁺ handling systems. The results collected suggest that the vasorelaxant effect of the CAM-68 alkaloid is associated with inhibition of NCX activity in reverse mode. It is well known that the KCl-induced contraction in blood vessel smooth muscle is based on the activation of voltage-dependent L-type Ca2+ channels and the influx of Ca2+ ions into the cell via membrane depolarisation.25,26From this perspective, the significant attenuation of KCl-induced contraction by the alkaloid CAM-68 indicates that it primarily inhibits the influx of Ca²⁺ ions through voltage-dependent Ca²⁺ channels. Such effects are characteristic of many natural alkaloids and are important in explaining their antihypertensive and vasodilator properties.27
The contractile response of vascular smooth muscle cells is closely linked to the dynamic changes in intracellular Ca²⁺ levels, and this factor is the primary determinant in the regulation of vascular tone.28,29The inhibitory effect of the CAM-68 alkaloid on CaCl₂ concentration-dependent contractions has provided a deeper insight into its mechanism of action. In isolated tissue models, contractions induced by altering the external Ca²⁺ concentration are primarily dependent on Ca²⁺ flux through the membrane, and this method is widely used in the evaluation of Ca²⁺ channel modulators.30,31In studies, the contractions induced by the cumulative addition of CaCl₂ in the absence of Ca²⁺ ions were effectively reduced by the alkaloid CAM-68. In the presence of pre-incubation with CAM-68, the decrease in contraction amplitude despite an increase in the external Ca²⁺ concentration indicates that it directly inhibits Ca²⁺ influx. This confirms that the CAM-68 alkaloid exerts a functional effect on voltage-dependent L-type Ca²⁺ channels. Moreover, we further confirmed this result by comparing it with verapamil, a specific blocker of L-type Ca²⁺ channels. One of the interesting aspects is that even under conditions of prior blockade with verapamil, the additional relaxant effect of the CAM-68 was retained. If the effect of CAM-68 were limited solely to the blockade of L-type Ca²⁺ channels, such an additional effect should not have been observed. Thus, alternative ion transport mechanisms or intracellular signaling routes might play a role in the vasorelaxant impact of the alkaloid CAM-68. If the effect of CAM-68 were limited solely to the blockade of L-type Ca²⁺ channels, such an additional effect should not have been observed. Thus, alternative ion transport mechanisms or intracellular signaling routes might play a role in the vasorelaxant impact of the alkaloid CAM-68. This additional relaxation strongly supports the alternative hypothesis that CAM-68 targets multiple pathways parallel to voltage-gated calcium channels, potentially cross-talking with receptor-operated systems or intracellular storage release.
In smooth muscle cells, Ca²⁺ homeostasis is one of the primary physiological processes and is of crucial importance for maintaining and regulating vascular tone.32This is primarily achieved via receptor-operated Ca2+ channels, which control the influx of Ca2+ through the membrane and operate in equilibrium with the sarcoplasmic reticulum.33,34In experiments, the alkaloid СAM-68 exhibited a dose-dependent relaxing effect on PE-induced aortic contractions. The aortic contraction induced by PE may be mediated through the activation of L-type Ca²⁺ channels and receptor-operated Ca²⁺ channels. Above, we mentioned that the CAM-68 alkaloid exerts a vasorelaxant effect by inhibiting L-type Ca²⁺ channels. Subsequent experiments were carried out in the presence of the α-adrenergic receptor blocker phentolamine to determine the involvement of receptor-operated Ca²⁺ channels.35In experiments involving phentolamine, the vasorelaxant effect of СAM-68 was observed to be reduced compared with the control.Thus, we can state that the receptor-operated Ca2+ channels participate in the vasorelaxant action of the alkaloid СAM-68 This can be explained by the reduced potency of the СAM-68 alkaloid when α₁-adrenergic receptors are blocked. This observation is consistent with established literature demonstrating that alpha-adrenergic receptor antagonists partially occlude the signaling pathways coupled to receptor-operated calcium channels, thereby limiting the dynamic range of multi-target vasodilators. Consequently, the distinct shift in CAM-68’s relaxant efficacy under phentolamine blockade provides robust functional evidence that its mechanism of action inherently involves the modulation of receptor-operated calcium influx parallel to voltage-gated pathways.
NCX in the blood vessels is an important transmembrane protein that regulates the intracellular Ca²⁺ concentration.36It operates in two main directions: it transports Ca²⁺ ions outwards in the forward direction and inwards in the reverse direction.37-39In a Krebs solution devoid of Na⁺ ions, slow-developing contractions in aortic rings promote the reverse-mode operation of the NCX, transporting Ca²⁺ into the cell and driving contraction.41Under these conditions, the CAM-68 (150 μM) significantly reduced the contraction force, indicating that the alkaloid inhibits Ca²⁺ influx via NCX. The specific NCX blocker KB-R7943 attenuates the vasorelaxant effect of the alkaloid CAM-68, further confirming that this pathway is primarily responsible.Moreover, in the presence of the Na⁺/K⁺-ATPase inhibitor ouabain, the rise in intracellular Na⁺ concentration drives the NCХ to operate in reverse mode, transporting Ca²⁺ ions into the cell.40,41In experiments, it was found that incubation of aortic rings with the alkaloid CAM-68 attenuated the contractions that developed under ouabain-incubation conditions. Under the conditions prevailing for KB-R7943, the alkaloid’s effect on the contractions that produce ouabain is significantly reduced. These results once again confirm that the vasorelaxant effect of the CAM-68 alkaloid is mediated via the NCX and is associated with inhibition of its reverse-mode activity. Collectively, the present results are in agreement with published evidence on the role of reverse-mode NCX in regulating vascular smooth muscle Ca²⁺ homeostasis and provide experimental support for the conclusion that suppression of NCX-dependent Ca²⁺ influx is one of the key mechanisms underlying the vasorelaxant action of CAM-68.
Conclusions
The CAM-68 alkaloid exerts its vasorelaxant effect on rat aortic rings through multiple complex mechanisms. CAM-68 inhibits the activity of voltage-dependent L-type Ca2+ channels and receptor-operated Ca2+ channels, and also attenuates the reverse-reversal mode of NCX. These effects are explained by modulation of intracellular Ca²⁺ homeostasis and inhibition of Ca²⁺ influx across the membrane.The vasorelaxant mechanism of action of the alkaloid CAM-68 is realised through a series of ion transport systems, manifesting as an important modulator of vascular tone regulation and potentially regarded as a pharmacologically promising bioactive compound.
Acknowledgement
The author would like to express sincere gratitude to the Laboratory of Cell Biophysics, Institute of Biophysics and Biochemistry at the National University of Uzbekistan, for providing the opportunity to conduct this research, as well as for the excellent laboratory equipment and technical support.
Funding Sources
This work was supported by the Ministry of Higher Education, Science and Innovation of Uzbekistan (grant AL-9224104346).
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
All experimental procedures involving laboratory animals were conducted in accordance with ethical guidelines approved by the Ethical Commission of the IBB at the National University of Uzbekistan.(This is documented under Protocol No. 7, dated 04/07/2022, regarding the utilization of research subjects). Segments of the aorta were isolated and prepared employing a previously described technique.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable.
Author Contributions
- SardorBakhtiyor ugli Sobirov: Methodology, Writing – review & editing, Writing – Original Draft,Resources, Data curation.
- AbdisalimAbdikarimovich Zaripov: Conceptualization, Methodology, Writing – review & editing,Writing – Original Draft,
- InoyatZulfiqorovich Zhumaev: Methodology, Writing – review & editing, Writing – original draft, Validation, Formal analysis.
- PulatBekmuratovich Usmanov: Conceptualization,Methodology, Writing – review & editing, Supervision, Resources, Funding acquisition, Project Administration.
- AdilbayTlepovich Esimbetov: Data Collection, Analysis, Writing – Review & Editing.
- ShavkatYusubovich Rustamov: Visualization,
- SadriddinNurillo ugli Boboev: Visualization,
- Gulnaz Sultanbayevna Begdullaeva: Data Collection, Analysis, Writing – Review & Editing.
- Eldor Bakhtiyor ugli Ibragimov: Resources, Data curation.
- Shakhnoza Bakhtiyorovna Qurbonova: Resources, Data curation.
- Sadoqat Abdukhalil qizi Safarova: Isolation of alkaloid CAM-68.
- Sherzod Niyatkobilovich Zhurakulov: Isolation of alkaloid CAM-68.
- Dilbar Djamalovna Safarova: Investigation, Formal analysis.
References
- Mills KT, Stefanescu A, He J. The global epidemiology of hypertension. Nat Rev Nephrol. 2020;16(4):223-237. DOI: 1038/s41581-019-0244-2.
CrossRef - Popa IP, Clim A, Pînzariu AC, Lazăr CI, Popa Ș, Tudorancea IM, Moscalu M, Șerban DN, Șerban IL, Costache-Enache II, Tudorancea I. Arterial Hypertension: Novel Pharmacological Targets and Future Perspectives. J Clin Med. 2024;13(19):5927. DOI: 3390/jcm13195927.
CrossRef - Schinzari F, Montenero R, Cardillo C, Tesauro M. Emerging Pharmacological Approaches for the Treatment of Arterial Hypertension. 2025;13(4):790. DOI10.3390/biomedicines13040790.
CrossRef - Suzuki Y, Giles WR, Zamponi GW, Kondo R, Imaizumi Y, Yamamura H. Ca²⁺ signaling in vascular smooth muscle and endothelial cells in blood vessel remodeling: a review. Inflamm Regen. 2024;44:1–14. DOI: 1186/s41232-024-00363-0.
CrossRef - Koide T, Giles WR, Kondo R, et al. Ca²⁺ microdomain-based excitation-transcription coupling in cardiac myocytes and vascular smooth muscle cells. Inflamm Regen. 2025;45:1–11. DOI:1186/s41232-025-00384-3.
CrossRef - Brozovich FV, Nicholson CJ, Degen CV, et al. Mechanisms of vascular smooth muscle contraction and the basis for pharmacologic treatment of smooth muscle disorders. Pharmacol Rev. 2016;68(2):476–532. DOI: 1124/pr.115.010652.
CrossRef - Pereira da Silva EA, Martín-Aragón Baudel M, Navedo MF, Nieves-Cintrón M. Ion channel molecular complexes in vascular smooth muscle. Front Physiol. 2022;13:1–10. DOI: 3389/fphys.2022.999369.
CrossRef - Dai C, Khalil RA. Calcium signaling dynamics in vascular cells and their deregulation in vascular disease. 2025;15(6):1–51. DOI: 10.3390/biom15060892
CrossRef - Melaku L, Elias B. The Physiological Mechanism of Extracellular Calcium-Sensing Receptor Action in the Regulation of Vascular Tone and Blood Pressure. Biomed J Sci Tech Res. 2023;51(5):43156-43165. DOI: 26717/BJSTR.2023.51.008172
CrossRef - Amssayef A, Eddouks M. Alkaloids as vasodilator agents: a review. Curr Pharm Des. 2023;29(24):1886–1895. DOI: 2174/1381612829666230809094313
CrossRef - Arash S, Mustafa G, Behzad Z. Neuropharmacological Potential of Diterpenoid Alkaloids. Pharmaceuticals. 2023;16(5):747. doi: 3390/ph16050747
CrossRef - Zaripov AA, Zhumaev IZ, Usmanov PB, et al. Mechanism of vasorelaxant action of isoquinoline alkaloid F-19 on rat aorta. Biomed Pharmacol J. 2025;18(2):1580–1589. DOI: https://dx.doi.org/10.13005/bpj/3195
CrossRef - Zhumaev IZ, Boboev SN, Usmanov PB, et al. Mechanism of positive inotropic effect of vincamine on cardiac muscle contraction activity. Biomed Pharmacol J. 2022;15:2309–2316. DOI: https://dx.doi.org/10.13005/bpj/2569
CrossRef - Thorneloe KS, Nelson MT. Ion channels in smooth muscle: regulators of intracellular calcium and contractility. Can J Physiol Pharmacol. 2005;83(3):215–242. DOI: 1139/y05-016
CrossRef - Pott C, Eckardt L, Goldhaber JI. Triple threat: the Na+/Ca2+ exchanger in the pathophysiology of cardiac arrhythmia, ischemia and heart failure. Curr Drug Targets. 2011 May;12(5):737-47. DOI: 2174/138945011795378559.
CrossRef - Sanders KM. Signal transduction in smooth muscle: mechanisms of calcium handling. J Appl Physiol (1985). 2001;91(3):1438–1449. DOI: 1152/jappl.2001.91.3.1438.
CrossRef - Gabbasov TM, Andrianova EI, Petrova SF, et al. Hydrolysis of lappaconitine in acidic solution: preparation of N-desacetyllappaconitin. Chem Nat Compd. 2020;56:767. DOI:1007/s10600-025-04857-2.
CrossRef - Kirill PC, Arkadiy OB, Dmitry SB. Synthesis, Pharmacological Evaluation, and Molecular Modeling of Lappaconitine–1,5-Benzodiazepine Hybrids. Molecules. 2023; 28(10):4234. doi: 3390/molecules28104234
CrossRef - Vandier C, Le Guennec JY, Bedfer G. What are the signaling pathways used by norepinephrine to contract the artery? Adv Physiol Educ. 2002;26:195–203. DOI: 1152/advan.00062.2001.
CrossRef - Gonzales RJ, Carter RW, Kanagy NL. Laboratory demonstration of vascular smooth muscle using rat aortic ring segments. Adv Physiol Educ. 2000;24:13–21.DOI: 1152/advances.2000.24.1.13.
CrossRef - Mirzayeva YT, Zaripov AA, Zhumaev IZ, et al. Protective effect of indole alkaloid vincanine against hypoxia-induced vasorelaxation in rat aorta. Biomed Pharmacol J. 2024;17(1):483–491. DOI: https://dx.doi.org/10.13005/bpj/2876.
CrossRef - Lee CH, Poburko D, Sahota P, et al. The mechanism of phenylephrine-mediated [Ca²⁺]i oscillations underlying tonic contraction. J Physiol. 2001;534(3):641–650. DOI: 1111/j.1469-7793.2001.t01-1-00641.x
CrossRef - Shattock M, Ottolia M, Bers DM, et al. Na⁺/Ca²⁺ exchange and Na⁺/K⁺-ATPase in the heart. J Physiol. 2015;593(6):1361–1381. DOI: 1113/jphysiol.2014.282319.
CrossRef - Ibragimov EB, Zhumaev IZ, Zaripov AA, et al. Mechanism of vasorelaxant effect of 12-hydroxynorfluorocurarine hydrochloride. J Appl Pharm Sci. 2026;16(3):283–291. DOI: 7324/JAPS.2026.272383.
CrossRef - Mironova GY, Haghbin N, Welsh DG. Functional tuning of vascular L-type Ca²⁺ channels. Front Physiol. 2022;13:1–6. DOI: 3389/fphys.2022.1058744.
CrossRef - Zaripov AA, Jumayev IZ, Usmanov PB, et al. Protective effect of DHQ-11 against hypoxia-induced vasorelaxation. Trends Sci. 2024;21(11):1–12.DOI: https://doi.org/10.48048/tis.2024.8192.
CrossRef - Triggle DJ. Calcium channel antagonists: clinical uses—past, present and future. Biochem Pharmacol. 2007;74(1):1–9. DOI: 1016/j.bcp.2007.01.016.
CrossRef - Amberg GC, Navedo MF. Calcium dynamics in vascular smooth muscle. 2013;20(4):281–289. DOI: 10.1111/micc.12046
CrossRef - Nathan RT, Erika MB, William FJ. Smooth Muscle Ion Channels and Regulation of Vascular Tone in Resistance Arteries and Arterioles. Compr Physiol. 2017; 7(2): p. 485–581. doi: 1002/cphy.c160011
CrossRef - Ghosh D, Syed AU, Prada MP, et al. Calcium channels in vascular smooth muscle. Adv Pharmacol. 2017;78:49–87. DOI: 1016/bs.apha.2016.08.002.
CrossRef - Khushmatov SS, Zhumaev IZ, Zhurakulov SN, Saidov AS, Vinogradova VI. Synthesis and comparative inotropic effects of isoquinoline alkaloids. Pharm Chem J. 2020;54:7–11. DOI 1007/s11094-020-02148-4.
CrossRef - Hu XQ, Zhang L. Role of transient receptor potential channels in the regulation of vascular tone. Drug Discov Today. 2024;29(7):104051. DOI: 1016/j.drudis.2024.104051.
CrossRef - McFadzean I, Gibson A. The developing relationship between receptor-operated and store-operated calcium channels. Br J Pharmacol. 2002;135(1):1–13. DOI: 1038/sj.bjp.0704468.
CrossRef - Jumayev IZ, Boboev SN, Usmanov PB, et al. Role of RyR2 and SERCA2a in cardioprotective effects. Trends Sci. 2025;22(1):1–11. DOI: https://doi.org/10.48048/tis.2025.8626
CrossRef - Saeed M, Sommer O, Holtz J, Bassenge E. Alpha-adrenoceptor blockade and beta-adrenergic vasodilation. J Cardiovasc Pharmacol. 1982;4(1):44–52. DOI: 1097/00005344-198201000-00008
CrossRef - Zhang S, Yuan JX, Barrett KE, Dong H. Role of Na⁺/Ca²⁺ exchange in regulating cytosolic Ca²⁺. Am J Physiol Cell Physiol. 2005;288(2):C245–C252. DOI: 1152/ajpcell.00411.2004.
CrossRef - Tykocki NR, Jackson WF, Watts SW. Reverse-mode Na⁺/Ca²⁺ exchange in venous contraction. Pharmacol Res. 2012;66(6):544–554. DOI: 1016/j.phrs.2012.08.004.
CrossRef - Qurbonova SB, Zhumaev IZ, Boboev SN, et al. Antiarrhythmic effect of isoquinoline derivative. Biomed Pharmacol J. 2025;18(3):2463–2473. DOI: https://dx.doi.org/10.13005/bpj/3270.
CrossRef - Raina H, Ella SR, Hill MA. Decreased activity of the smooth muscle Na⁺/Ca²⁺ exchanger. J Physiol. 2008;586(6):1669–1681. DOI: 1113/jphysiol.2007.150268.
CrossRef - Orth T, Pyanova A, Lux S, et al. Vascular smooth muscle BK channels limit vasoconstriction. FASEB J. 2024;38(17):1–16. DOI:1096/fj.202400628RR.
CrossRef - Staehr C, Aalkjaer C, Matchkov VV. The vascular Na,K-ATPase: clinical implications. Clin Sci (Lond). 2023;137(20):1595–1618. DOI: 1042/CS20220796.
CrossRef

