
Manuscript accepted on :31-12-2025
Published online on: 21-01-2026
Plagiarism Check: Yes
Reviewed by: Dr Nasir Abdelrafie
Second Review by: Dr. Exbrayat Jean-Marie
Final Approval by: Dr. Mariia Shanaida
Aryo Tedjo1,2* , Novi Yantih3, Fadilah2and Mohamad Rafi4
, Novi Yantih3, Fadilah2and Mohamad Rafi4
1Department of Medical Chemistry, Faculty of Medicine, Universitas Indonesia, Jalan Salemba Raya 6, Jakarta, Indonesia
2Doctoral Program in Pharmaceutical Sciences, Universitas Pancasila, Jalan Srengseng sawah, Jakarta, Indonesia
3Faculty of Pharmacy, Universitas Pancasila, Jalan Srengseng sawah, Jakarta, Indonesia
4Department of Chemistry, Faculty of Mathematics and Natural Sciences, IPB University, JalanTanjung Kampus IPB Dramaga, Bogor, Indonesia
Corresponding Author E-mail: aryo.tedjo@ui.ac.id
DOI : https://dx.doi.org/10.13005/bpj/3352
Abstract
Breast cancer of the luminal subtype remains a major cause of morbidity and mortality among women worldwide, largely due to the development of resistance to endocrine therapies such as tamoxifen. One of the transcription factors involved in the proliferation of breast cancer cells is E2F2, whose activity is regulated by the CDK4/6–Rb complex. This study aimed to evaluate the potential of active compounds in the 75% ethanol extract of Annona muricata (soursop) leaves as CDK6 inhibitors and suppressors of E2F2 expression in MCF-7 breast cancer cells. LC–MS/MS analysis identified 152 unique compounds, most of which belonged to the acetogenin class. Activity prediction using SkelSpheres descriptors based on support vector regression (SVR) yielded an R²-testing value of 0.8925, indicating high predictive validity for CDK6 inhibitory activity. Several compounds, including annonacin, muricin D, and longifolicin, exhibited predicted IC₅₀ values below 1 µM. Molecular docking results showed strong affinity between acetogenins and CDK6, with rerank scores comparable to the reference ligand ribociclib. In vitro cytotoxicity testing demonstrated that the extract had an IC₅₀ value of 9.7 ppm against MCF-7 cells. Gene expression analysis using RT-qPCR revealed a significant reduction in E2F2 expression following extract treatment (0.5175 ± 0.1732) compared with the control group (1.0087 ± 0.1517; p = 0.000), comparable to the effect of tamoxifen. These findings suggest that acetogenins present in the 75% ethanol extract of A. muricata leaves possess potential as anticancer candidates by suppressing E2F2 expression, presumably through the inhibition of CDK6.
Keywords
Annona muricata; CDK6 inhibitor; E2F2 expression; In silico; In vitro; MCF-7 breast cancer
Download this article as:| Copy the following to cite this article: Tedjo A, Yantih N, Fadilah F, Rafi M. Acetogenin-Rich Annona muricata Leaf Extract as a CDK6 Inhibitor and E2F2 Suppressor in MCF-7 Breast Cancer Cells. Biomed Pharmacol J 2026;19(1). |
| Copy the following to cite this URL: Tedjo A, Yantih N, Fadilah F, Rafi M. Acetogenin-Rich Annona muricata Leaf Extract as a CDK6 Inhibitor and E2F2 Suppressor in MCF-7 Breast Cancer Cells. Biomed Pharmacol J 2026;19(1).Available from: https://bit.ly/3Lt3rAd |
Introduction
Breast cancer is one of the leading causes of morbidity and mortality among women worldwide. According to recent global cancer statistics, breast cancer is the most frequently diagnosed cancer and the leading cause of cancer-related death among women. Together with lung and colorectal cancer, it accounts for about 51% of all new cancer diagnoses in women, with breast cancer alone contributing approximately 32% of cases.1 The luminal A and luminal B subtypes generally exhibit dependence on estrogen signaling to sustain cellular proliferation and survival. One of the primary therapeutic strategies for luminal breast cancer is endocrine therapy, such as tamoxifen, which functions by blocking the binding of estrogen to its receptor and thereby suppressing the transcription of estrogen-regulated genes. However, Resistance to tamoxifen represents a major challenge in endocrine therapy.2 Such resistance is frequently associated with overexpression of EGFR and activation of downstream molecules within alternative signaling pathways, including ERK1/2, PI3K, and AKT.3 Consequently, molecular approaches are required to identify novel genetic targets involved in the mechanisms of resistance and sensitivity to this therapy.2,3
One of the transcription factors that plays a critical role in breast cancer progression is E2F2, which regulates the G1/S transition through the activation of genes controlling DNA replication and cell proliferation. Several studies have reported that E2F2 is overexpressed in breast cancer tissues compared with normal tissues, contributing to enhanced cell proliferation and resistance to apoptosis.4–6 The activity of E2F2 is regulated by the CCND1–CDK4/6–Rb pathway, in which the CCND1–CDK4/6 complex phosphorylates the Rb protein, leading to the release of E2F2, which subsequently activates the transcription of proliferative genes.7 Therapeutically, CDK4/6 inhibitors such as palbociclib, ribociclib, and abemaciclib have been shown to prevent Rb phosphorylation, retain E2F2 in its inhibited form, and slow tumor growth.7 Thus, CDK6 has emerged as a key target worthy of further evaluation as a major regulator within the breast cancer proliferation pathway.
A variety of bioactive compounds derived from plants have been reported to modulate the expression of genes involved in cellular malignancy, as demonstrated by phytochemicals such as sulforaphane from broccoli that regulate cell cycle progression, apoptosis, and enzyme activity.8 Similarly, the leaves of Annona muricata (soursop) contain bioactive compounds with the potential to modulate cancer-related gene expression and thus represent a promising source of anticancer agents. Compounds such as annonacin have been shown to influence the regulation of apoptosis-related genes, including ER, cyclin D1, and BCL2, in MCF-7 breast cancer cells.9 In addition, species of the genus Annona have been widely reported to possess anticancer properties, supported by ethnomedicinal use and diverse secondary metabolites such as acetogenins and alkaloids with documented antiproliferative activity.10 Another compound, astragalin (kaempferol-3-O-β-D-glucoside), exhibits cytotoxic activity against M.D. Anderson – Metastatic Breast 231 (MDA-MB-231) breast cancer cells, and based on in silico analyses has potential interactions with proteins such as BCL-2, CDK2, CDK4, MAPK, and RAF1.11
This study aimed to identify the compounds present in the 75% ethanol extract of A. muricata leaves using LC–MS/MS analysis and to determine which of these compounds have the potential to suppress E2F2 expression, particularly through the inhibition of CDK6 as a key upstream regulator. By integrating metabolite profiling, in silico prediction of CDK6 inhibitory activity, molecular docking, and in vitro assays in MCF-7 breast cancer cells, this research seeks to elucidate the molecular mechanisms through which A. muricata exerts its anticancer effects. The findings are expected to provide new insights into natural product–based modulation of the CDK6–Rb–E2F2 axis and to contribute to the development of alternative therapeutic candidates for overcoming endocrine resistance in luminal breast cancer.
Materials and Methods
Extraction of Simplicia and Identification of Chemical Constituents of A. muricata Leaf Extract
A total of 20 g of Annona muricata leaf simplicia was dried to a low moisture content and ground into a fine powder. Extraction was performed using the maceration method with 75% ethanol (1:10 w/v) for 24 hours with intermittent stirring, followed by filtration using Whatman filter paper. The procedure was repeated once to ensure optimal extraction, after which the combined filtrates were evaporated using a rotary evaporator at 50°C to obtain a viscous extract. A total of 15 mg of the extract was reconstituted to a final concentration of 1500 ppm. An aliquot of 200 µL was vortexed, dried using a nitrogen evaporator, and stored at –80°C prior to analysis.
Chemical component analysis was conducted using LC–MS/MS on a Vanquish Ultra-High Performance Liquid Chromatography (UHPLC) system coupled to a Q Exactive Plus Orbitrap HRMS (Thermo Scientific). Separation was achieved on an Accucore C18 column (100 × 2.1 mm; 1.5 µm) at 30°C using eluent A (H₂O + 0.1% formic acid) and eluent B (acetonitrile + 0.1% formic acid). The gradient program consisted of 5–30% B (0–12.5 min), increasing to 95% B (33 min), and returning to 5% B (40 min), with a flow rate of 0.2 mL/min and an injection volume of 2 µL. Mass spectrometric analysis was performed in the m/z range of 100–1500 under positive ionization mode to obtain the metabolite profile.
Prediction of the Activity of 75% Ethanolic A. muricata Leaf Extract Compounds as CDK6 Inhibitors
Prediction of the inhibitory potential (IC50) of compounds identified from the 75% ethanolic extract of Annona muricata leaves against CDK6 was conducted following the method of Suryandari et al.12 The IC₅₀ prediction model was developed using bioactivity data retrieved from ChEMBL via DataWarrior v6.1.0, comprising in vitro-tested drug-like compounds.13,14 Active and inactive compounds were clustered using SkelSpheres descriptors with a similarity threshold of 90%, which evaluates structural similarity based on circular atom fragments.13 One representative compound from each cluster was designated as training data, while the remaining compounds were used as test data in a SVR model. The model was considered valid when the correlation between actual and predicted pIC₅₀ values yielded R²-testing > 0.515, after which it was used to estimate the IC₅₀ values of metabolites detected in the A. muricata extract.
Activity Confirmation by Molecular Docking
Molecular docking analysis was performed using Molegro Virtual Docker (MVD v7.0.0) to analyze the interaction between the compound with the lowest predicted IC₅₀ and CDK6 protein. The crystal structure of CDK6 (PDB ID: 5L2T) was obtained from the Research Collaboratory for Structural Bioinformatics – Protein Data Bank (RCSB PDB) and prepared by removing water molecules and correcting amino acid residues.16 The protein and ligand structures were further prepared by assigning partial atomic charges and adding explicit hydrogens to ensure chemical completeness. Ligand geometry was optimized using the “Ligand Energy Inspector” tool, a built-in module in Molegro Virtual Docker used for energy minimization and conformational optimization of ligands prior to docking.17 Docking validity was assessed based on Root Mean Square Deviation (RMSD) < 2 Å using 20 independent runs. Energy parameters including MolDock Score, Re-rank Score, and H-bond interactions were evaluated to determine binding affinity, with the Re-rank Score used as the primary indicator.17
Cytotoxicity Assay Using the MTT Method
MCF-7 breast cancer cells (5 × 10³ cells/100 µL) were cultured in 96-well microplates using Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS) and penicillin–streptomycin. The cells were treated with the 75% ethanolic extract of A. muricata leaves at eight concentrations (1–1000 ppm), along with a negative control, for 24 hours under conditions of 37°C and 5% CO₂. After incubation, MTT solution (5 mg/mL) was added and the cells were further incubated for 6 hours. The reaction was terminated with 10% Sodium Dodecyl Sulfate (SDS) and left overnight at room temperature in the dark. Formazan intensity was measured using an Enzyme-Linked Immunosorbent Assay (ELISA) reader at 595 nm to determine cell viability. The IC₅₀ value was calculated from the concentration–response curve generated from triplicate experiments.
Measurement of E2F2 Expression by Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
Gene expression analysis was performed by extracting total RNA from MCF-7 cells treated with the extract at a concentration corresponding to 1/10 × IC₅₀ from the MTT assay. A total of 200 ng of purified RNA was used as a template for cDNA synthesis via reverse transcription. Quantification of E2F2 gene expression was carried out using real-time quantitative PCR (RT-qPCR). The primer pair used consisted of Forward (5′–GAGAAGACTCGGTATGACAC–3′) and Reverse (5′–TGGGGTCTTCAAACATTCC–3′). The annealing temperature was determined based on a melting curve analysis indicating the most specific amplification without primer-dimer formation.
Relative gene expression was normalized to the housekeeping gene ACTB and calculated using the Livak and Schmittgen (2⁻ΔΔCt) method, where ΔCt represents the difference between the target gene and ACTB Ct values. ΔΔCt represents the difference between treated and control samples.18 Relative quantification (R) was calculated using the equation R = 2(-ΔΔCt), where:
ΔΔCt = ΔCt_sample − ΔCt_control = (Ct_target − Ct_HK)_sample − (Ct_target −Ct_HK)_control
as described by Pfaffl et al.19 The Ct (cycle threshold) value represents the number of cycles required for the fluorescence signal to cross the detection threshold. Statistical analysis using the t-test was performed to compare mean gene expression levels between the treatment groups, namely plant extract and tamoxifen, and the control group.
Results
Extraction of A. muricata Leaf Simplicia and Identification of Secondary Metabolites
The secondary metabolite profile of the 75% ethanolic extract of A. muricata leaves was analyzed using LC–MS/MS in positive electrospray ionization mode (ESI⁺). The total ion chromatogram (TIC) displayed a complex elution pattern with several major peaks detected across the 0–40 min retention time range (Figure 1). Peaks with relatively high intensities were observed at RT 2.1, 8.4, 12.9, 14.0, 28.7, and 34.8 minutes. The most intense peak appeared at RT 14.0 minutes and was identified as the base peak, indicating that the compound eluting at this time was the major constituent of the extract. The distribution of peaks across the chromatographic window suggests that 75% ethanol efficiently extracts compounds with a broad polarity range, including polar to semi-nonpolar constituents.
A total of 858 ion features (m/z) or distinct mass spectra were detected in the LC–MS/MS chromatogram. Among these, 334 features were successfully annotated by matching mass and fragmentation patterns with the instrument database and PubChem. After removing duplicate entries, 152 unique compounds were obtained. These 152 compounds were subsequently subjected to activity prediction as CDK6 inhibitors.
Prediction of CDK6 Inhibitory Activity of Metabolites Detected in the 75% Ethanolic Extract of A. muricata
The validation results of the CDK6 inhibitor activity prediction model developed using DataWarrior with SkelSpheres descriptors and SVR are presented in Figure 2. Figure 2 shows a strong correlation between actual and predicted pIC₅₀ values. The model yielded a coefficient of determination (R²-testing) of 0.8925, indicating that approximately 89.25% of the variance in actual pIC₅₀ values could be explained by the model. The clustering of data points near the linear regression line demonstrates high consistency between experimental and predicted activity, confirming that the combination of SkelSpheres descriptors and SVR provides strong predictive performance in modeling the QSAR relationship of CDK6 inhibitors. Predicted activities of the compounds detected in the 75% ethanolic extract of A. muricata leaves against CDK6 are shown in Table 1. All identified metabolites fell within the “excellent” inhibitor category (IC₅₀ < 1 µM).17
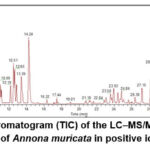 |
Figure 1: Total ion chromatogram (TIC) of the LC–MS/MS analysis of the 75% ethanolic leaf extract of Annona muricata in positive ionization mode (ESI⁺). |
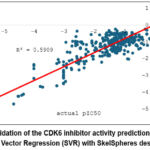 |
Figure 2: Validation of the CDK6 inhibitor activity prediction model using Support Vector Regression (SVR) with SkelSpheres descriptors. |
Confirmation of Predicted Activity by Molecular Docking
Confirmation of metabolite activity as CDK6 inhibitors, as listed in Table 1, was performed using molecular docking. Validation of the docking procedure using the native ligand ribociclib on the CDK6 receptor (PDB ID: 6ZZ9) is presented in Table 2. Docking results demonstrated consistent and valid outcomes. The MolDock score ranged from –165.487 to –178.379 kJ/mol, with rerank scores between –128.438 and –142.612 kJ/mol. The best pose was obtained from the conformation with the lowest MolDock score (–178.379 kJ/mol) and an RMSD of 1.474 Å, which is below the 2 Å threshold, indicating accurate reproduction of the ligand’s crystallographic orientation. Hydrogen bond energy values ranged from –4.038 to –6.819 kJ/mol, reflecting stable hydrogen interactions between ribociclib and key active site residues of CDK6.
Table 1: Predicted activity of compounds detected in the 75% ethanolic extract of A. muricata leaves as CDK6 inhibitors.
| CID | Compound | RT (min) | IC50 prediction (µM) | Activity Criteria* |
| 10897073 | muricin D | 26.397 | 0.00721 | excellent |
| 132942 | bullatalicin | 27.889 | 0.00725 | excellent |
| 5742590 | daucosterol | 33.584 | 0.00727 | excellent |
| 10722577 | asiminecin | 30.905 | 0.00731 | excellent |
| 44584143 | muricatocin A | 22.418 | 0.00734 | excellent |
| 44584476 | annoreticuin | 30.269 | 0.00734 | excellent |
| 14456327 | annonacin | 27.251 | 0.00734 | excellent |
| 11093061 | cis-corossolone | 24.726 | 0.00737 | excellent |
| 11028548 | cis-annomontacin | 30.941 | 0.00740 | excellent |
| 177313 | longifolicin | 30.475 | 0.00741 | excellent |
*Indriyanto et al.20
Table 2: Validation of the molecular docking protocol using redocking of the native ligand ribociclib into the CDK6 receptor (PDB ID: 6ZZ9).
| Ligand Pose | Ligand Name | MolDock Score (kJ/mol) | Rerank Score (kJ/mol) | Hbond (kJ/mol) | RMSD (Å) |
| [01]6ZZ_900 [A] | ribociclib | -177.763 | -142.612 | -6.819 | 0.766 |
| [00]6ZZ_900 [A] | ribociclib | -178.379 | -140.979 | -6.031 | 1.474 |
| [02]6ZZ_900 [A] | ribociclib | -165.487 | -128.438 | -4.038 | 1.557 |
Molecular docking results for the metabolites identified in the 75% ethanolic extract of A. muricata leaves are summarized in Table 3. Table 3 shows variations in binding affinity values, reflecting differing inhibitory potentials. The reference ligand ribociclib displayed a MolDock score of –175.652 kJ/mol, rerank score of –132.609 kJ/mol, and hydrogen bond energy of –5.994 kJ/mol, and was used as the benchmark for high CDK6 affinity. Several acetogenins—such as annonacin (–150.757 kJ/mol), cis-corossolone (–127.926 kJ/mol), and asiminecin (–123.460 kJ/mol)—exhibited MolDock scores relatively close to ribociclib, suggesting strong interactions with the active site residues of CDK6. However, compounds with high torsion numbers (>25), such as muricatocin A and bullatalicin, showed markedly increased rerank scores (21.508–88.959 kJ/mol). Ligands with high flexibility may undergo significant conformational adjustments to optimize nonbonded interactions, resulting in internal energy penalties that reduce overall docking stability.21
Figure 3 illustrates the interactions between CDK6 and two ligands, namely the native ligand (6ZZ_900 [A]/ribociclib) and the test ligand (longifolicin). The three-dimensional protein structure is shown with α-helices in red, β-sheets in blue, and loops in gray. Ligands occupying the binding pocket are displayed as stick models, colored green for the native ligand and yellow for the test ligand. Panel (a) shows ribociclib interacting with crucial residues such as Asp104, Ile19, Val101, Leu152, and Asp163 through hydrogen bonding and hydrophobic interactions. Panel (b) demonstrates that longifolicin also engages with several active-site residues, including Ile19, Gln149, Glu21, Lys147, Gly25, and Asp163. These comparisons indicate that longifolicin is capable of mimicking key interactions of the native ligand within the CDK6 active site, suggesting its potential as an effective CDK6 inhibitor.
Table 3: Docking results of metabolites identified in the 75% ethanolic extract of A. muricata leaves as CDK6 inhibitors.
| Ligand Pose | MolDock Score (kJ/mol) | Rerank Score (kJ/mol) | Torsions | Hbond (kJ/mol) |
| [00]6zz_900 [a] (ribociclib) | -175.652 | -132.609 | 5 | -5.994 |
| [00]longifolicin | -119.304 | -69.492 | 26 | -4.175 |
| [00]annonacin | -150.757 | -42.997 | 26 | -3.058 |
| [02]annoreticuin | -92.994 | -39.271 | 26 | -2.500 |
| [06]daucosterol | -95.418 | -38.132 | 9 | -3.266 |
| [07]muricin d | -90.976 | -36.478 | 24 | -6.555 |
| [01]cis-annomontacin | -95.838 | -23.274 | 28 | -2.104 |
| [00]cis-corossolone | -127.926 | 10.802 | 26 | -4.721 |
| [02]muricatocin a | -110.521 | 21.508 | 26 | -1.476 |
| [00]asiminecin | -123.460 | 27.402 | 25 | -4.775 |
| [00]bullatalicin | -121.178 | 88.959 | 25 | -0.461 |
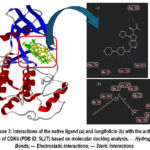 |
Figure 3: Interactions of the native ligand (a) and longifolicin (b) with the active site of CDK6 (PDB ID: 5L2T) based on molecular docking analysis. — Hydrogen Bonds; — Electrostatic Interactions; — Steric Interactions |
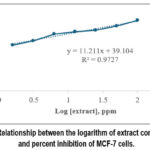 |
Figure 4: Relationship between the logarithm of extract concentration and percent inhibition of MCF-7 cells. |
Cytotoxicity Assay Using the MTT Method
The cytotoxic activity of the 75% ethanolic extract of Annona muricata against MCF-7 breast cancer cells using the MTT assay is shown in Figure 4. The extract exhibited concentration-dependent inhibition, with increasing extract concentrations corresponding to higher percentages of growth inhibition. Linear regression analysis yielded the equation y = 11.211x + 39.104 with an R² value of 0.9727, indicating a strong correlation between the logarithm of extract concentration and percent inhibition. Based on the regression model, the IC₅₀ value, defined as the concentration producing 50% inhibition, was calculated to be 9.3735 ppm.
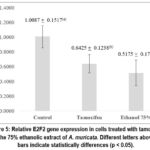 |
Figure 5: Relative E2F2 gene expression in cells treated with tamoxifen and the 75% ethanolic extract of A. muricata. Different letters above the bars indicate statistically differences (p < 0.05). Click here to view Figure |
Measurement of E2F2 Expression Using Reverse Transcription Quantitative PCR (RT-qPCR)
Relative expression of the E2F2 gene normalized to the housekeeping gene ACTB is presented in Figure 5. Different letters above the expression bars indicate statistically significant differences among treatments (p < 0.05). Figure 5 shows significant differences between treatment groups and the control. The control group exhibited the highest expression level of 1.0087 ± 0.1517, which differed significantly from the 75% ethanolic extract group (0.5175 ± 0.1732; p = 0.000) and the tamoxifen group (0.6425 ± 0.1238; p = 0.001). Meanwhile, no significant difference was observed between the extract and tamoxifen (p = 0.149). These results indicate that both tamoxifen and the 75% ethanolic extract of A. muricata significantly reduced E2F2 gene expression relative to the control, with comparable degrees of downregulation. This suggests that acetogenins in the extract possess potential CDK6 inhibitory activity as predicted in silico, supported by significant cytotoxic effects on MCF-7 cells and reduced E2F2 gene expression.
Discussion
The predictive model for CDK6 inhibitors constructed in this study yielded an R²-testing value of 0.8925, indicating that the model meets and even surpasses the minimum threshold for strong external predictive performance. According to established QSAR model validation criteria, a model is considered to possess good predictive ability when it fulfills statistical parameters such as R² > 0.6, cross-validation (R²-CV) > 0.5, or R²-testing > 0.5.22,23 The predicted activities of compounds detected in the 75% ethanol extract of A. muricata leaves through LC–MS/MS analysis against the CDK6 proteon showed excellent inhibitory potential based on their IC₅₀ values, as all identified compounds fell into the “excellent” inhibitor category (IC₅₀ < 1 µM).20 Most of these compounds are secondary metabolites of the acetogenin class, including muricin D, bullatalicin, asiminecin, muricatocin A, annoreticuin, annonacin, cis-corossolone, cis-annomontacin, and longifolicin, which are widely known for their cytotoxic and antiproliferative activities against various cancer cell types.24,25 The very low predicted IC₅₀ values indicate that the acetogenin constituents of soursop extract have strong potential to interact with the active site of CDK6, a key regulator of the G1–S phase transition in the cell cycle.
Molecular docking results further demonstrated that the compounds identified as “excellent” inhibitors exhibit favorable interactions with the inhibitory site of the CDK6 protein. Acetogenins such as muricin D and annoreticuin displayed negative rerank scores accompanied by stable hydrogen-bond interactions, reinforcing the hypothesis that most acetogenins in A. muricata extract function as CDK6 inhibitors through strong binding affinity at the enzyme’s active site. Likewise, compounds including muricin D and annoreticuin consistently showed negative rerank scores with stable hydrogen interactions, supporting the notion that these acetogenins may serve as potent CDK6 inhibitors by engaging the catalytic domain of the protein.
The cytotoxicity assay confirmed that the 75% ethanol extract of A. muricata possesses strong cytotoxic activity against MCF-7 cells, indicating its potential as an anticancer candidate. These findings are in line with those of Tianing et al.26, who reported that a 96% ethanol extract of A. muricata leaves also exhibited cytotoxic effects against 4T1 breast cancer cells. Measurement of E2F2 expression in MCF-7 cells treated with the extract at 1/10 × IC₅₀ revealed a reduction in E2F2 gene expression. This decrease, which was comparable to the effect of tamoxifen, suggests that the extract may suppress proliferative pathways through modulation of the CDK4/6–Rb–E2F2 axis or a crosstalk mechanism that ultimately influences this transcription factor. Nevertheless, because the current experiments employed crude extract and the in silico results remain predictive, further studies are needed, including the isolation of active acetogenins (e.g., annonacin, muricin D, longifolicin) and experimental validation of CDK6 inhibition, Rb phosphorylation status, and E2F2 protein expression, to confirm the molecular mechanisms and therapeutic potential of this extract more specifically.
Conclusion
The 75% ethanol extract of Annona muricata leaves exhibited strong cytotoxic activity against MCF-7 breast cancer cells, with an IC₅₀ value of 9.37 ppm. Treatment with the extract also significantly reduced E2F2 gene expression compared with the control, with a level of suppression comparable to that of tamoxifen. In silico analysis of compounds identified by LC–MS/MS indicated that most active constituents—particularly acetogenins such as annonacin, muricin D, longifolicin, and annoreticuin—possess strong potential as CDK6 inhibitors, with predicted IC₅₀ values < 1 µM and binding affinities approaching that of the reference ligand ribociclib. These findings provide evidence that acetogenins present in the 75% ethanol extract of A. muricata leaves may suppress cancer cell proliferative activity, and the observed reduction in E2F2 expression is likely mediated through CDK6 inhibition.
Acknowledgement
We acknowledge the Department of Medical Chemistry, Faculty of Medicine, Universitas Indonesia to facilitate our work.
Funding Sources
The authors are thankful to the Ministry of Higher Education, Science, and Technology of the Republic of Indonesia for funding this reaseach through the 2025 Regular Fundamental Grant Program No. SP DIPA-139.04.1.69332
Conflict of Interest
The author(s) do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
- Aryo Tedjo : Data Collection, Methodology, Analysis, Writing – Original Draft.
- Novi Yantih: Methodology, Supervision, Project Administration, Funding Acquisition
- Fadilah: Conceptualization, Supervision, Methodology, Review & Editing
- Mohamad Rafi: Conceptualization, Supervision
References
- Siegel RL, Kratzer TB, Giaquinto AN, et al. Cancer statistics, 2025. CA Cancer J Clin. 2025;75(1):10-45.
CrossRef - Rondón-Lagos M, Villegas VE, Rangel N, et al. Tamoxifen Resistance: Emerging Molecular Targets. Int J Mol Sci. 2016;17(8):1357-1388.
CrossRef - Riggins RB, Schrecengost RS, Guerrero MS, et al. Pathways to tamoxifen resistance. Cancer Lett. 2007;256(1):1-24. doi: 10.1016/j.canlet.2007.03.016.
CrossRef - Li Y, Huang J, Yang D, et al. Expression patterns of E2F transcription factors and their potential prognostic roles in breast cancer. Oncol Lett. 2018;15(6):9216-9230.
CrossRef - Liu ZL, Bi XW, Liu PP, et al. Expressions and prognostic values of the E2F transcription factors in human breast carcinoma. Cancer Manag Res. 2018;10:3521-3532.
CrossRef - Gao Y, Qiao X, Liu Z, et al. The role of E2F2 in cancer progression and its value as a therapeutic target. Front Immunol. 2024;15:1397303-1397311.
CrossRef - Spring L, Bardia A. Cycling Toward Progress: Ribociclib, a CDK 4/6 Inhibitor for Breast Cancer. Clin Cancer Res. 2018;24(13):2981-2983.
CrossRef - Gasmi A, Gasmi Benahmed A, Shanaida M, et al. Anticancer activity of broccoli, its organosulfur and polyphenolic compounds. Crit Rev Food Sci Nutr. 2024;64(22):8054-8072.
CrossRef - Ko YM, Wu TY, Wu YC, et al. Annonacin induces cell cycle-dependent growth arrest and apoptosis in estrogen receptor-α-related pathways in MCF-7 cells. J Ethnopharmacol. 2011;137(3):1283–12
CrossRef - Nugraha AS, Damayanti YD, Wangchuk P, et al. Anti-Infective and Anti-Cancer Properties of the Annona Species: Their Ethnomedicinal Uses, Alkaloid Diversity, and Pharmacological Activities. Molecules. 2019;24(23):4419-4450.
CrossRef - Araujo-Padilla X, Ramón-Gallegos E, Díaz-Cedillo F, et al. Astragalin identification in graviola pericarp indicates a possible participation in the anticancer activity of pericarp crude extracts: In vitro and in silico approaches. Arabian Journal of Chemistry. 2022;15(4):103720-103729.
CrossRef - Suryandari DA, Tedjo A, Fadilah F. Differentially Expressed Genes (DEGs) Analysis and In Silico Studies Identify Tumor Necrosis Factor (TNF) Inhibition and Peroxisome Proliferator-Activated Receptor Alpha (PPARA) Activation as Targets for Gallic Acid Derivatives in Insulin Resistance. Trop J Nat Prod Res. 2024;8(12):9476 – 9485.
CrossRef - Sander T, Freyss J, von Korff M, et al. DataWarrior: an open-source program for chemistry aware data visualization and analysis. J Chem Inf Model. 2015;55:460–473.
CrossRef - Mendez D, Gaulton A, Bento AP, et al. ChEMBL: towards direct deposition of bioassay data. Nucleic Acids Res. 2019;47:D930–D940.
CrossRef - OECD (2014), Guidance Document on the Validation of (Quantitative) Structure-Activity Relationship [(Q)SAR] Models, OECD Series on Testing and Assessment, No. 69, OECD Publishing, Paris
- Berman HM, Westbrook J, Feng Z, et al. The Protein Data Bank. Nucleic Acids Res. 2000;28:235-242.
CrossRef - Molexus molegro virtual docker user manual. P. Baatrupsvej 41, DK-8300 Odder, Denmark: 2019
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. 2001;25(4):402-408
CrossRef - Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002;30(9):e36-e46.
CrossRef - Indrayanto G, Putra GS, Suhud F. Validation of in-vitro bioassay methods: Application in herbal drug research. Profiles Drug Subst Excip Relat Methodol. 2021;46:273-307
CrossRef - Peach ML, Cachau RE, Nicklaus MC. Conformational energy range of ligands in protein crystal structures: The difficult quest for accurate understanding. J Mol Recognit. 2017;30(8):1-32.
CrossRef - Golbraikh A, Tropsha A. Predictive QSAR modeling based on diversity sampling of experimental datasets for the training and test set selection. J Comput Aided Mol Des. 2002;16(5-6):357-369. 10.1023/a:1020869118689.
CrossRef - Pratim Roy P, Paul S, Mitra I, et al. On two novel parameters for validation of predictive QSAR models. Molecules. 2009;14(5):1660-1 doi.10.3390/molecules14051660.
CrossRef - Yuan SS, Chang HL, Chen HW, et al. Annonacin, a mono-tetrahydrofuran acetogenin, arrests cancer cells at the G1 phase and causes cytotoxicity in a Bax- and caspase-3-related pathway. Life Sci. 2003:9;72(25):2853-2861. 10.1016/s0024-3205(03)00190-5.
CrossRef - Liaw CC, Chang FR, Lin CY, et al. New cytotoxic monotetrahydrofuran annonaceous acetogenins from Annona muricata. J Nat Prod. 2002;65(4):470-475.10.1021/np0105578.
CrossRef - Tianing NW, Sumardika IW, Wiryanthini IAD, et al. In Vitro Evaluation of Soursop Leaf (Annona muricata L) Ethanol Extract against Development and Proliferation of 4T1 Breast Cancer Cell. Trop J Nat Prod Res. 2025;9(3):921–924. doi.org/10.26538/tjnpr/v9i3.5.
CrossRef
List of Abbreviations
ACTB – Beta-Actin
AKT – Protein Kinase B
BCL2 – B-cell Lymphoma 2
CCND1 – Cyclin D1
CDK4/6 – Cyclin-Dependent Kinase 4/6
CDK6 – Cyclin-Dependent Kinase 6
E2F2 – E2F Transcription Factor 2
EGFR – Epidermal Growth Factor Receptor
ER – Estrogen Receptor
ERK1/2 – Extracellular Signal-Regulated Kinase 1 and 2
LC-MS/MS – Liquid Chromatography–Tandem Mass Spectrometry
MAPK – Mitogen-Activated Protein Kinase
MCF-7 – Michigan Cancer Foundation-7 (garis sel kanker payudara luminal A)
MTT – 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl Tetrazolium Bromide
PI3K– Phosphoinositide 3-Kinase
RAF1 – Rapidly Accelerated Fibrosarcoma 1
Rb – Retinoblastoma Protein

