
Manuscript accepted on :08-06-2026
Published online on: 16-06-2026
Plagiarism Check: Yes
Reviewed by: Dr. Rashid Axundov
Second Review by: Dr. Akmal El-Mazny
Final Approval by: Dr. Patorn Piromchai
Shakhnoza Bakirova3* , Shahzod Ergashev2, Sirojiddin Omonturdiyev1,3, Izzatullo Abdullaev1, Ulugbek Gayibov1, Shakhlokhon Ismailova4, Kamila Eshbakova2,Mirtaza Allamuratov3
, Shahzod Ergashev2, Sirojiddin Omonturdiyev1,3, Izzatullo Abdullaev1, Ulugbek Gayibov1, Shakhlokhon Ismailova4, Kamila Eshbakova2,Mirtaza Allamuratov3
1Department of Plantcytoprotectors, Institute of Bioorganic Chemistry named after A.S.Sadykov, Tashkent, Uzbekistan
2Department of Terpenoids and Phenolic Compounds Laboratory, Institute of the Chemistry of Plant Substances named after Academician S. Yu. Yunusov, Uzbekistan
3Departmentof Humanphysiology, National University of Uzbekistan named after Mirzo Ulugbek, Tashkent, Uzbekistan
4Deparment of Human physiology, Namangan State University, Uzbekistan
Corresponding Author E-mail: shahnozabositxonova@gmail.com
Abstract
Intracellular calcium homeostasis plays a crucial role in the regulation of vascular smooth muscle contraction and relaxation. The present study investigated the vasorelaxant activity of ZS 22-12 using isolated rat thoracic aorta and in silico molecular docking analysis of calcium-regulating proteins. ZS 22-12 significantly inhibited both KCl-induced and phenylephrine-induced contractions in a concentration-dependent manner, with IC₅₀ values of 38.23 μM and 43.57 μM, respectively. Comparative studies with verapamil and phentolamine suggested that the vasorelaxant effect of ZS 22-12 is associated with modulation of voltage-dependent and receptor-operated calcium signaling pathways. To clarify the molecular mechanism, docking studies were performed with the Ca²⁺ L-type channel, Ca²⁺ R-type channel, SERCA, ryanodine receptor (RyR), Na⁺/Ca²⁺ exchanger (NCX), and Ca²⁺-ATPase. ZS 22-12 demonstrated stable binding affinities toward all investigated targets through hydrophobic, hydrogen bonding, and electrostatic interactions. Overall, the combined in vitro and in silico results indicate that ZS 22-12 acts as a multitarget modulator of calcium homeostasis in vascular smooth muscle cells and exhibits promising vasorelaxant potential.
Keywords
Calcium channels; Molecular docking; NCX; Ryanodine receptor; SERCA; Vascular Smooth muscle; Vasorelaxant activity; ZS 22-12
| Copy the following to cite this article: Bakirova S, Ergashev S, Omonturdiyev S, Abdullaev I, Gayibov U, Ismailova S, Eshbakova K, Allamuratov M. From Molecular Docking to Vascular Ion Channel Mechanisms: A Multitarget Study of ZS 22-12. Biomed Pharmacol J 2026;19(2). |
| Copy the following to cite this URL: Bakirova S, Ergashev S, Omonturdiyev S, Abdullaev I, Gayibov U, Ismailova S, Eshbakova K, Allamuratov M. From Molecular Docking to Vascular Ion Channel Mechanisms: A Multitarget Study of ZS 22-12. Biomed Pharmacol J 2026;19(2). Available from: https://bit.ly/43C0yTa |
Introduction
Cardiovascular diseases remain one of the leading causes of morbidity and mortality worldwide and are closely associated with disturbances in vascular tone regulation, endothelial dysfunction, oxidative stress, and abnormal calcium homeostasis.1,2 Contraction and relaxation of vascular smooth muscle cells are highly dependent on the precise regulation of intracellular Ca²⁺ concentration, which is controlled by a complex network of membrane-associated and intracellular calcium transport systems. Dysregulation of these pathways contributes to the development of hypertension, vasospasm, ischemic injury, and other vascular disorders.3,4,5
Among the major calcium-regulating mechanisms, voltage-dependent L-type Ca²⁺ channels play a fundamental role in mediating extracellular calcium influx during membrane depolarization. Activation of these channels increases intracellular calcium concentration and promotes smooth muscle contraction.6,7 In addition, receptor-operated calcium signaling pathways activated through G protein-coupled receptors (GPCRs), particularly α₁-adrenergic receptors, also contribute significantly to vascular contractility by stimulating intracellular calcium mobilization through phospholipase C-dependent pathways.8,9 Furthermore, intracellular calcium transport proteins such as sarco/endoplasmic reticulum Ca²⁺-ATPase (SERCA), ryanodine receptors (RyR), Na⁺/Ca²⁺ exchangers (NCX), and Ca²⁺-ATPases are critically involved in maintaining calcium balance by regulating calcium release, reuptake, and extrusion processes within vascular smooth muscle cells.10,11
Currently used calcium channel blockers, including verapamil and other clinically available vasodilators, are effective in the treatment of hypertension and cardiovascular disorders; however, long-term administration may be associated with adverse effects and limited selectivity. Therefore, identification of novel biologically active compounds capable of modulating multiple calcium-associated signaling pathways remains an important area of pharmacological research.
In recent years, increasing attention has been directed toward the investigation of synthetic and naturally derived compounds with vasorelaxant and calcium-regulating properties.12,13 Many of these compounds exert their biological effects through interactions with ion channels, membrane transport proteins, and intracellular calcium-handling systems. Understanding the molecular mechanisms underlying these interactions is essential for the development of new therapeutic agents targeting vascular dysfunction.14,15
ZS 22-12 is a biologically active compound with potential calcium-modulating activity; however, its effects on vascular smooth muscle calcium transport systems have not yet been fully elucidated. Therefore, the present study aimed to investigate the vasorelaxant effects of ZS 22-12 on isolated rat thoracic aorta and to evaluate its possible mechanisms of action through both in vitro pharmacological experiments and in silico molecular docking analysis.16,17 The effects of the compound were assessed using KCl-induced and phenylephrine-induced contraction models to determine its influence on voltage-dependent and receptor-operated calcium pathways. In addition, molecular docking studies were performed to evaluate the interaction of ZS 22-12 with several calcium-regulating proteins, including L-type and R-type calcium channels, SERCA, RyR, NCX, and Ca²⁺-ATPase. The combined experimental and computational approach was employed to clarify the potential role of ZS 22-12 as a multitarget regulator of calcium homeostasis in vascular smooth muscle cells.18,19
Materials and Methods
Animal Ethics
All experimental protocols and surgical manipulations involving animals were reviewed and approved by the Institutional Animal Care and Use Committee before commencement of the study. Experimental animals were maintained in the institutional vivarium under standardized environmental conditions, including a temperature of 22 ± 2 °C and relative humidity of 55–65%, with unrestricted access to standard pellet diet and drinking water. All procedures involving laboratory animals were performed in accordance with the European Directive 2010/63/EU concerning the protection of animals used for scientific purposes. Ethical approval for the study was provided by the Animal Ethics Committee of the Institute of Bioorganic Chemistry, Academy of Sciences of the Republic of Uzbekistan (Protocol No. 133/1a/h).
Molecular Docking
Computational investigations were performed using freely available academic software packages. Three-dimensional structures of calcium-associated target proteins were retrieved from the Protein Data Bank (PDB), an international repository containing experimentally determined biomolecular structures [7–9]. The selected targets included the voltage-gated L-type calcium channel Cav1.2 (PDB ID: 6JP5), sodium/calcium exchanger NCX1 (PDB ID: 8SGI), ryanodine receptor type 2 (RyR2; PDB ID: 5C33), and sarcoplasmic/endoplasmic reticulum Ca²⁺-ATPase (SERCA; PDB ID: 6RB2).
The structures of ZS 22-12 and reference ligands were initially generated in two-dimensional format using ChemDraw (PerkinElmer, USA). The compounds were then transformed into three-dimensional conformations and energy-minimized in Avogadro software employing the MMFF94 force field to obtain stable molecular geometries. Additional information regarding molecular descriptors and physicochemical properties was obtained from the PubChem database.20,21
Visualization and structural examination of proteins and ligand–protein complexes were carried out using PyMOL (version 1.2). Docking simulations were conducted using AutoDock 4.2 (The Scripps Research Institute), whereas docking grids and parameter files were prepared using AutoDock Tools (ADT). This molecular docking approach allows prediction of ligand orientation, binding affinity, and interaction patterns based on flexible ligand and rigid receptor models, thereby facilitating interpretation of ligand–target interactions.22,23
Tissue Preparation
All experimental procedures involving animals were performed according to internationally recognized recommendations for laboratory animal care and were fully consistent with the European Directive 2010/63/EU regarding the use of animals for scientific purposes.24,25 Animals were housed under controlled vivarium conditions at 22 ± 2 °C and 55–65% humidity with ad libitum access to food and water.
Thoracic aortas were isolated from adult male Wistar rats weighing 200–250 g. Prior to tissue collection, animals were anesthetized with sodium pentobarbital to reduce pain and stress. Following anesthesia, euthanasia was performed by cervical dislocation. The thoracic cavity was then opened, and the thoracic aorta was carefully excised and immediately placed into a 5 mL organ bath containing Krebs–Henseleit physiological solution. The composition of the solution (mM) was as follows: NaCl 120.4, KCl 5.0, NaHCO₃ 15.5, NaH₂PO₄ 1.2, MgCl₂ 1.2, CaCl₂ 2.5, glucose 11.5, and HEPES buffer adjusted to pH 7.4, as commonly applied in vascular reactivity studies. In some experiments, Ca²⁺-free Krebs solution containing EGTA (1 mM) was used to investigate calcium-dependent responses.26,27
All physiological solutions were continuously oxygenated with carbogen gas (95% O₂ and 5% CO₂) and maintained at 37 °C using a thermostatically regulated water bath. Connective and adipose tissues surrounding the vessels were carefully removed, after which the aorta was sectioned into ring preparations measuring approximately 3–4 mm in length.
Aortic Ring Contraction Studies
Aortic ring segments were mounted on platinum hooks and attached to an isometric force transducer system (Radnoti, USA) for recording vascular contractile activity. The tissues were allowed to equilibrate for 60 min under an initial resting tension of 1 g (10 mN), which is considered a standard preload for rat thoracic aorta experiments. Changes in isometric force were amplified and digitally recorded using an automated acquisition system. Contractile responses were measured in millinewtons (mN) and subsequently expressed as percentages of maximal contraction for comparative statistical analysis.28,29
Statistical Analysis
Experimental data are expressed as mean ± standard deviation (SD) derived from at least three independent experiments (n = 3-6). Statistical differences between multiple groups were analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc multiple comparison test. Comparisons between two experimental groups were evaluated using the unpaired Student’s t-test. Statistical significance was accepted at p < 0.05. Graphical presentation of the data, including bar charts, concentration–response curves, and error bars, was performed using GraphPad Prism and OriginPro 2022a (OriginLab, USA). All statistical procedures were conducted according to standard biomedical research practices.
Results
Effect of ZS 22-12 on voltage-dependent and receptor-operated Ca²⁺ channels in isolated rat aorta
The involvement of Ca²⁺ transport systems in the vasorelaxant activity of ZS 22-12 was investigated using isolated rat thoracic aorta preparations precontracted with KCl and phenylephrine. High extracellular KCl induces membrane depolarization and activation of voltage-dependent L-type Ca²⁺ channels; therefore, this model was used to evaluate the effect of ZS 22-12 on voltage-operated calcium influx. In parallel, phenylephrine-induced contraction was employed to assess receptor-operated Ca²⁺ channels associated with α₁-adrenergic GPCR signaling.30,31
As presented in Figure 1A), ZS 22-12 caused a concentration-dependent inhibition of KCl-induced vascular contraction. At concentrations of 10, 30, 40, 60, and 70 μM, the contractile force decreased to 89.57 ± 1.85%, 72.90 ± 2.18%, 55.03 ± 1.98%, 25.04 ± 2.60%, and 22.07 ± 2.24%, respectively. The calculated IC₅₀ value for ZS 22-12 in the KCl-induced contraction model was 38.23 μM. These data indicate that the compound effectively suppresses depolarization-mediated Ca²⁺ influx through L-type calcium channels.
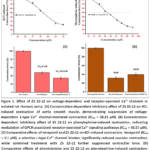 |
Figure 1: Effect of ZS 22-12 on voltage-dependent and receptor-operated Ca²⁺ channels in isolated rat thoracic aorta. |
To further clarify the mechanism of action, the effect of ZS 22-12 was compared with verapamil, a classical L-type Ca²⁺ channel blocker. As shown in Figure 1C, KCl-induced contraction in the control group was maintained at 100 ± 1.1%, whereas verapamil reduced the contractile response to 48 ± 2.4%. Combined treatment with verapamil and ZS 22-12 further decreased contraction to 35 ± 3.2%. Verapamil exhibited a markedly lower IC₅₀ value (0.1 μM) compared with ZS 22-12 (38.23 μM), indicating the stronger potency of the reference blocker. Nevertheless, the similar inhibitory profile observed for both compounds suggests that ZS 22-12 exerts its vasorelaxant activity, at least partially, via modulation of voltage-dependent L-type Ca²⁺ channels.32,33
The participation of receptor-operated Ca²⁺ channels in the activity of ZS 22-12 was assessed using phenylephrine-induced contraction. Phenylephrine activates α₁-adrenergic receptors coupled with GPCR-mediated phospholipase C signaling, resulting in intracellular Ca²⁺ mobilization and vascular smooth muscle contraction. As illustrated in Figure 1B, ZS 22-12 significantly inhibited phenylephrine-induced contraction in a concentration-dependent manner. At concentrations of 10, 30, 40, 50, 62, 70, 80, and 90 μM, the contractile force values were reduced to 85.2 ± 2.09%, 70.0 ± 1.80%, 53.0 ± 2.38%, 45.0 ± 1.86%, 32.0 ± 1.87%, 25.0 ± 2.30%, 18.0 ± 2.60%, and 15.0 ± 2.40%, respectively. The calculated IC₅₀ value was 43.57 μM.
To confirm the involvement of α₁-adrenergic receptor-mediated signaling pathways, the effects of ZS 22-12 were compared with phentolamine, a non-selective α-adrenergic receptor antagonist. As shown in Figure 1D, phenylephrine-induced contraction in the control group reached 100 ± 1.1%, while phentolamine reduced the contraction force to 32 ± 3.2%. In the presence of phentolamine and ZS 22-12, the contractile response was 40 ± 3.4%. These findings indicate that ZS 22-12 interferes with GPCR-dependent calcium signaling pathways involved in vascular smooth muscle contraction.34,35
Collectively, the obtained results demonstrate that ZS 22-12 possesses significant vasorelaxant activity associated with both voltage-dependent L-type Ca²⁺ channels and receptor-operated calcium transport systems. The inhibitory effects observed in KCl- and phenylephrine-induced contraction models suggest that the compound modulates extracellular Ca²⁺ influx as well as receptor-mediated intracellular calcium mobilization in vascular smooth muscle cells.
Molecular docking analysis of ZS 22-12 with Ca²⁺ L-type ion channel (PDB ID: 6JP5)
Based on the in vitro pharmacological experiments, ZS 22-12 demonstrated a pronounced inhibitory effect on KCl-induced contraction of isolated rat aortic smooth muscle. Since KCl-mediated depolarization activates voltage-dependent L-type Ca²⁺ channels and promotes extracellular calcium influx into vascular smooth muscle cells, the obtained experimental results suggested that the vasorelaxant activity of ZS 22-12 may be associated with modulation of these channels. Therefore, to clarify the possible molecular mechanism of action and confirm the affinity of the compound toward voltage-dependent calcium transport systems, an in silico molecular docking study was performed using the crystal structure of the Ca²⁺ L-type ion channel (PDB ID: 6JP5).
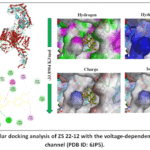 |
Figure 2: Molecular docking analysis of ZS 22-12 with the voltage-dependent Ca²⁺ L-type ion channel (PDB ID: 6JP5). |
The docking analysis revealed a favorable binding affinity of ZS 22-12 toward the Ca²⁺ L-type ion channel, with a calculated binding energy of −33.8904 kJ/mol, indicating the formation of a stable ligand–receptor complex. Visualization of the docking pose demonstrated that the ligand occupied a deeply localized region within the hydrophobic binding cavity of the channel, suggesting potential modulation of calcium ion transport activity.
Two-dimensional interaction analysis demonstrated that ZS 22-12 interacted with several amino acid residues within the binding pocket, including VAL226, ASP236, LEU237, TYR238, ARG242, ARG243, PRO244, ASP454, ALA455, GLU457, PRO553, SER555, and GLN556. Hydrophobic interactions were mainly observed with VAL226, LEU237, PRO244, and ARG242, which may contribute significantly to stabilization of the ligand inside the channel cavity. Additionally, polar and electrostatic interactions involving TYR238, ARG243, ASP454, SER555, and GLN556 may further enhance binding specificity and receptor affinity (Figure 2).
Surface interaction analysis revealed that hydrophobic interactions constituted the predominant stabilizing force in the ligand–protein complex. Hydrophobicity mapping showed that ZS 22-12 occupied a predominantly nonpolar region of the receptor cavity, whereas hydrogen bond surface analysis indicated the presence of moderate polar contacts around the ligand-binding region. Charge distribution analysis demonstrated interactions with both positively and negatively charged residues, supporting electrostatic stabilization of the complex. Furthermore, ionizability surface mapping suggested that the ligand was accommodated within a chemically favorable microenvironment suitable for stable complex formation.
Overall, the molecular docking findings are in agreement with the in vitro vascular experiments and support the hypothesis that ZS 22-12 exerts its vasorelaxant activity through direct interaction with voltage-dependent Ca²⁺ L-type ion channels. The stable binding profile and interactions with functionally important amino acid residues suggest that the compound may interfere with calcium influx in vascular smooth muscle cells, thereby contributing to suppression of KCl-induced contraction.
Molecular docking analysis of ZS 22-12 with Ca²⁺ R-type ion channel
In addition to voltage-dependent L-type calcium channels, receptor-associated calcium transport systems may also contribute to vascular smooth muscle contraction and regulation of intracellular Ca²⁺ homeostasis. Since the pharmacological experiments demonstrated that ZS 22-12 significantly inhibited phenylephrine-induced contraction mediated through GPCR-associated calcium signaling pathways, molecular docking analysis was further performed to evaluate the possible interaction of ZS 22-12 with the Ca²⁺ R-type ion channel.
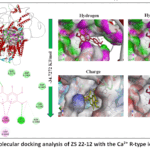 |
Figure 3: Molecular docking analysis of ZS 22-12 with the Ca²⁺ R-type ion channel. |
The docking analysis demonstrated that ZS 22-12 exhibited a stable interaction with the Ca²⁺ R-type ion channel, with a calculated binding energy of −34.7272 kJ/mol. The obtained binding affinity was slightly stronger than that observed for the Ca²⁺ L-type channel, suggesting favorable accommodation of the ligand within the receptor cavity. Three-dimensional visualization indicated that the ligand occupied a central hydrophobic region of the channel pocket, which may contribute to modulation of receptor-associated calcium influx mechanisms.
Two-dimensional interaction mapping revealed that ZS 22-12 interacted with several amino acid residues, including THR1338, TRP1341, THR1342, LYS1345, ASN1466, TYR1469, ARG1472, GLN1174, ASP1468, ASP1465, ASP1483, PRO1479, and HIS1480. Among these residues, hydrophobic and π-alkyl interactions were predominantly observed with PRO1479, TYR1469, and TRP1341, contributing to stabilization of the ligand within the binding cavity (Figure 3). In addition, hydrogen bond interactions involving ARG1472 and LYS1345 may strengthen ligand–protein affinity and enhance binding specificity. Electrostatic contacts with ASP1483 and ASP1468 further suggest the contribution of polar interactions to stabilization of the complex.36
Surface property analysis demonstrated that hydrophobic interactions constituted one of the major stabilizing forces in the ligand–receptor complex. Hydrophobicity mapping revealed that ZS 22-12 occupied a partially nonpolar region of the channel cavity, while hydrogen bond surface analysis indicated the presence of several polar donor and acceptor regions surrounding the ligand. Charge distribution analysis showed interactions with both negatively and positively charged residues within the active site environment. Furthermore, ionizability mapping demonstrated that the ligand was localized within a chemically compatible microenvironment favorable for stable complex formation.
Overall, the obtained docking results support the experimental findings and indicate that ZS 22-12 may modulate receptor-associated Ca²⁺ transport systems through stable interaction with the Ca²⁺ R-type ion channel. The favorable binding affinity and multiple hydrophobic, hydrogen bonding, and electrostatic interactions suggest that the compound may contribute to suppression of receptor-mediated calcium influx and vascular smooth muscle contraction.
Molecular docking analysis of ZS 22-12 with SERCA
Intracellular calcium homeostasis in vascular smooth muscle cells is tightly regulated not only by membrane-associated calcium channels but also by sarco/endoplasmic reticulum Ca²⁺-ATPase (SERCA), which plays a critical role in transporting cytosolic Ca²⁺ into the sarcoplasmic reticulum. Impairment or modulation of SERCA activity directly influences intracellular calcium accumulation, vascular tone, and smooth muscle relaxation. Since the in vitro experiments demonstrated significant vasorelaxant effects of ZS 22-12 associated with calcium transport systems, molecular docking analysis was performed to investigate the possible interaction of the compound with SERCA.37
 |
Figure 4: Molecular docking analysis of ZS 22-12 with sarco/endoplasmic reticulum Ca²⁺-ATPase (SERCA). |
The docking results demonstrated that ZS 22-12 formed a stable complex with SERCA, exhibiting a binding affinity of −31.38 kJ/mol. The ligand was localized within the hydrophobic region of the receptor cavity, suggesting favorable accommodation inside the active binding pocket of the protein. Although the calculated binding energy was slightly lower than those observed for the Ca²⁺ L-type and R-type channels, the obtained value still indicates thermodynamically favorable ligand–protein interaction.
Two-dimensional interaction analysis revealed that ZS 22-12 interacted with several amino acid residues, including ASP703, ARG560, PHE487, GLU442, MET494, and LYS515. Hydrophobic interactions were predominantly observed with PHE487 and MET494, which may contribute to stabilization of the ligand inside the receptor cavity. In addition, electrostatic interactions involving ARG560 and polar contacts with GLU442 and LYS515 may support ligand recognition and binding specificity. The interaction profile suggests that both hydrophobic and electrostatic forces contribute to stabilization of the ligand–SERCA complex (Figure 4).
Surface property analysis demonstrated that hydrophobicity represented one of the major stabilizing factors within the binding pocket. Hydrogen bond surface mapping indicated the presence of several donor and acceptor regions surrounding the ligand, whereas charge distribution analysis revealed interactions between ZS 22-12 and charged amino acid residues localized around the active site. Ionizability surface analysis further demonstrated that the ligand occupied a chemically compatible microenvironment favorable for stable complex formation.
Overall, the obtained docking results suggest that ZS 22-12 may interact with SERCA and potentially modulate intracellular calcium reuptake mechanisms in vascular smooth muscle cells. Together with the in vitro vascular findings, these results indicate that the vasorelaxant activity of ZS 22-12 may involve not only inhibition of membrane-associated calcium influx but also regulation of intracellular calcium homeostasis through interaction with SERCA.
Molecular docking analysis of ZS 22-12 with ryanodine receptor (RyR)
Ryanodine receptors (RyRs) are intracellular calcium-release channels localized on the sarcoplasmic reticulum membrane and play a central role in regulation of intracellular Ca²⁺ dynamics in vascular smooth muscle cells. Activation of RyR channels induces release of stored Ca²⁺ from intracellular calcium reservoirs into the cytoplasm, thereby contributing to muscle contraction and calcium-dependent signaling processes. Since the vasorelaxant effects of ZS 22-12 observed in vitro may involve modulation of intracellular calcium mobilization, molecular docking analysis was performed to evaluate the possible interaction of ZS 22-12 with the RyR channel.
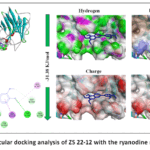 |
Figure 5: Molecular docking analysis of ZS 22-12 with the ryanodine receptor (RyR). |
The docking analysis demonstrated that ZS 22-12 formed a stable complex with RyR, exhibiting a binding affinity of −31.38 kJ/mol. The ligand was positioned within a hydrophobic cavity of the receptor structure, indicating favorable accommodation inside the active binding region of the channel. The obtained binding energy suggests thermodynamically stable ligand–protein interaction that may influence RyR-associated calcium release mechanisms.
Two-dimensional interaction analysis revealed that ZS 22-12 interacted with several amino acid residues, including GLU701, GLY708, GLY709, GLY710, GLU711, GLU712, TYR703, THR700, LEU787, and GLY786. Hydrophobic interactions were mainly observed with LEU787 and GLY710, contributing to stabilization of the ligand inside the receptor cavity. In addition, hydrogen bond and polar interactions involving THR700, GLU701, GLU711, and TYR703 may support ligand recognition and binding specificity (Figure 5). The interaction profile indicates that both hydrophobic and electrostatic interactions participate in stabilization of the ligand–RyR complex.38
Surface visualization analysis demonstrated that hydrophobicity represented one of the dominant stabilizing forces in the binding pocket. Hydrogen bond surface mapping showed the presence of several donor and acceptor regions surrounding the ligand-binding area, whereas charge distribution analysis revealed interactions between ZS 22-12 and charged amino acid residues localized around the active site. Ionizability surface analysis further indicated that the ligand occupied a chemically favorable microenvironment suitable for stable complex formation.
Overall, the obtained docking results suggest that ZS 22-12 may interact with RyR-associated intracellular calcium-release systems and potentially contribute to regulation of intracellular Ca²⁺ mobilization in vascular smooth muscle cells. Together with the in vitro pharmacological findings, these results support the hypothesis that the vasorelaxant activity of ZS 22-12 involves modulation of both extracellular calcium influx and intracellular calcium-release pathways.
3.X. Molecular docking analysis of ZS 22-12 with Na⁺/Ca²⁺ exchanger (NCX)
The Na⁺/Ca²⁺ exchanger (NCX) is one of the major calcium transport systems involved in regulation of intracellular Ca²⁺ homeostasis in vascular smooth muscle cells. NCX plays a critical role in maintaining cellular calcium balance by exchanging intracellular Ca²⁺ for extracellular Na⁺, thereby contributing to vascular contraction–relaxation processes. Since the in vitro experiments demonstrated that ZS 22-12 significantly modulated vascular smooth muscle contraction associated with calcium transport pathways, molecular docking analysis was performed to evaluate the possible interaction of ZS 22-12 with NCX.39
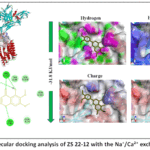 |
Figure 6: Molecular docking analysis of ZS 22-12 with the Na⁺/Ca²⁺ exchanger (NCX). |
The docking analysis demonstrated that ZS 22-12 formed a stable ligand–protein complex with NCX, exhibiting a binding affinity of −31.8 kJ/mol. The ligand was localized within the active hydrophobic cavity of the exchanger, indicating favorable accommodation inside the receptor binding region. The obtained binding energy suggests thermodynamically stable interaction that may influence NCX-mediated calcium transport activity.
Two-dimensional interaction analysis revealed that ZS 22-12 interacted with several amino acid residues, including ASN96, THR98, SER100, ASN101, ARG219, ASP218, TYR224, LYS225, LYS229, GLN74, GLU579, ASP577, and ILE576. Hydrogen bond interactions were mainly observed with ASN96, ARG219, and ASP218, suggesting stabilization of the ligand within the active site. Hydrophobic and π-alkyl interactions involving TYR224 and LYS225 may additionally contribute to stabilization of the ligand–protein complex. Furthermore, polar interactions with THR98, SER100, and ASN101 may support ligand recognition and binding specificity (Figure 6).
Surface property analysis demonstrated that hydrophobic interactions represented one of the dominant stabilizing forces within the binding cavity. Hydrogen bond surface mapping revealed the presence of multiple donor and acceptor regions surrounding the ligand-binding area. Charge distribution analysis indicated interactions between ZS 22-12 and both positively and negatively charged residues localized around the active site environment. In addition, ionizability surface analysis demonstrated that the ligand occupied a chemically compatible microenvironment favorable for stable complex formation.
Overall, the obtained docking results suggest that ZS 22-12 may interact with NCX-associated calcium transport systems and potentially contribute to modulation of intracellular calcium exchange mechanisms in vascular smooth muscle cells. Together with the in vitro pharmacological findings, these results support the hypothesis that the vasorelaxant activity of ZS 22-12 may involve regulation of both calcium influx and Na⁺/Ca²⁺ exchange-dependent intracellular calcium homeostasis.
Molecular docking analysis of ZS 22-12 with Ca²⁺-ATPase
Ca²⁺-ATPase plays a fundamental role in maintaining intracellular calcium homeostasis by actively transporting Ca²⁺ ions across cellular membranes through ATP-dependent mechanisms. Regulation of Ca²⁺-ATPase activity is essential for controlling cytosolic calcium concentration, vascular smooth muscle relaxation, and calcium-dependent signaling processes. Since the in vitro pharmacological studies demonstrated that ZS 22-12 significantly modulated calcium-associated contractile responses in isolated aortic smooth muscle, molecular docking analysis was performed to evaluate the possible interaction of ZS 22-12 with Ca²⁺-ATPase.
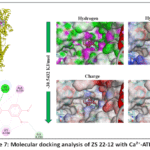 |
Figure 7: Molecular docking analysis of ZS 22-12 with Ca²⁺-ATPase. |
The docking analysis demonstrated that ZS 22-12 formed a stable ligand–protein complex with Ca²⁺-ATPase, exhibiting a binding affinity of −30.5432 kJ/mol. The ligand was localized within the hydrophobic region of the receptor cavity, suggesting favorable accommodation inside the active binding pocket of the protein. Although the obtained binding energy was slightly lower than those observed for voltage-dependent calcium channels and NCX, the interaction still indicates thermodynamically favorable complex formation.
Two-dimensional interaction analysis revealed that ZS 22-12 interacted with several amino acid residues, including GLU125, MET126, LYS158, GLY211, ALA724, and LYS727. Hydrogen bond interactions were predominantly observed with MET126 and LYS727, contributing to stabilization of the ligand within the active site. Hydrophobic and π-alkyl interactions involving ALA724 and LYS158 may additionally support stabilization of the ligand–protein complex. Electrostatic interaction with GLU125 further suggests the contribution of polar contacts to binding specificity and receptor affinity (Figure 7).
Surface property analysis demonstrated that hydrophobic interactions represented one of the major stabilizing forces within the receptor cavity. Hydrogen bond surface mapping indicated the presence of donor and acceptor regions surrounding the ligand-binding environment, while charge distribution analysis revealed interactions between ZS 22-12 and both positively and negatively charged residues localized around the active site. Furthermore, ionizability surface analysis demonstrated that the ligand occupied a chemically favorable microenvironment suitable for stable complex formation.
Overall, the obtained docking results suggest that ZS 22-12 may interact with Ca²⁺-ATPase-associated calcium transport systems and potentially contribute to regulation of intracellular calcium extrusion mechanisms in vascular smooth muscle cells. Together with the in vitro pharmacological findings, these results support the hypothesis that the vasorelaxant activity of ZS 22-12 involves modulation of multiple calcium-regulating transport pathways associated with vascular smooth muscle relaxation.40
Discussion
Regulation of intracellular Ca²⁺ homeostasis is one of the principal mechanisms controlling vascular smooth muscle contraction and relaxation. Disturbances in calcium transport systems contribute directly to the development of vascular dysfunction, hypertension, vasospasm, and oxidative stress-associated cardiovascular disorders. In the present study, the vasorelaxant activity of ZS 22-12 was evaluated using both in vitro pharmacological experiments and in silico molecular docking analyses targeting several calcium-regulating proteins associated with vascular smooth muscle function.
The obtained in vitro results demonstrated that ZS 22-12 significantly inhibited KCl-induced contraction of isolated rat thoracic aorta in a concentration-dependent manner. Since high extracellular KCl depolarizes the plasma membrane and activates voltage-dependent L-type Ca²⁺ channels, suppression of KCl-induced contraction strongly suggests inhibition of extracellular calcium influx through these channels. The calculated IC₅₀ value of ZS 22-12 in the KCl model (38.23 μM) further confirmed its pronounced inhibitory activity toward depolarization-mediated contractile responses. Comparison with verapamil, a classical L-type calcium channel blocker, revealed a similar inhibitory profile, supporting the hypothesis that ZS 22-12 may modulate voltage-operated calcium channels in vascular smooth muscle cells.
Phenylephrine-induced contraction experiments additionally demonstrated that ZS 22-12 significantly suppressed GPCR-associated contractile signaling pathways. Phenylephrine activates α₁-adrenergic receptors coupled with phospholipase C-mediated intracellular calcium mobilization and receptor-operated calcium entry. The observed concentration-dependent inhibition of phenylephrine-induced contraction by ZS 22-12, with an IC₅₀ value of 43.57 μM, indicates that the compound may interfere not only with voltage-dependent calcium influx but also with receptor-mediated intracellular calcium signaling pathways. Comparative analysis with phentolamine further supports the possible involvement of α₁-adrenergic receptor-associated calcium transport mechanisms in the vasorelaxant effect of the compound.
To further clarify the molecular basis of these pharmacological effects, molecular docking analyses were performed with several calcium-regulating proteins involved in vascular smooth muscle contraction. Docking studies demonstrated that ZS 22-12 exhibited stable binding affinities toward all investigated targets, including the Ca²⁺ L-type ion channel, Ca²⁺ R-type ion channel, SERCA, ryanodine receptor (RyR), Na⁺/Ca²⁺ exchanger (NCX), and Ca²⁺-ATPase. Among these targets, the strongest interaction was observed with the Ca²⁺ R-type ion channel (−34.7272 kJ/mol), followed by the Ca²⁺ L-type ion channel (−33.8904 kJ/mol), indicating a high affinity of ZS 22-12 toward calcium channel-associated transport systems. These findings are in good agreement with the experimental vascular studies demonstrating suppression of KCl- and phenylephrine-induced contractions.
The docking interaction profiles revealed that hydrophobic interactions represented one of the dominant stabilizing forces contributing to ligand–protein complex formation. In addition, hydrogen bonding, electrostatic interactions, and polar contacts with amino acid residues localized within the active cavities of the proteins contributed to stabilization and binding specificity of ZS 22-12. The surface analyses of hydrogen bonding, hydrophobicity, charge distribution, and ionizability further demonstrated that the ligand occupied chemically favorable microenvironments within the receptor cavities, supporting stable complex formation.
Importantly, the docking results suggest that the vasorelaxant activity of ZS 22-12 is likely mediated through a multitarget calcium-regulatory mechanism rather than interaction with a single calcium transport system. Interaction with voltage-dependent Ca²⁺ channels may suppress extracellular calcium influx, whereas modulation of SERCA, RyR, NCX, and Ca²⁺-ATPase may additionally influence intracellular calcium storage, release, extrusion, and exchange processes. Such combined regulation of intracellular calcium homeostasis may contribute to effective suppression of vascular smooth muscle contraction.
Despite the promising findings obtained in the present study, several limitations should be acknowledged. First, the vascular experiments were performed using isolated rat thoracic aorta preparations, which may not fully reflect the complexity of in vivo cardiovascular physiology and systemic hemodynamic regulation. Second, although molecular docking analyses demonstrated stable interactions between ZS 22-12 and multiple calcium-regulating proteins, docking studies alone cannot fully confirm direct molecular mechanisms or functional modulation of these targets under physiological conditions. Additional electrophysiological investigations, calcium imaging studies, and molecular biology approaches are required to validate the precise mechanisms of action of ZS 22-12 on specific calcium transport systems. Furthermore, the present study did not evaluate long-term toxicity, pharmacokinetic properties, or in vivo antihypertensive efficacy of the compound, which are essential for future therapeutic development. Therefore, further comprehensive experimental and preclinical studies are necessary to confirm the pharmacological potential and clinical relevance of ZS 22-12 as a multitarget regulator of vascular calcium homeostasis.
Taken together, the present findings demonstrate that ZS 22-12 possesses significant vasorelaxant activity associated with modulation of both membrane-associated and intracellular calcium transport systems. The combined in vitro and in silico results indicate that ZS 22-12 may act as a multitarget regulator of calcium homeostasis in vascular smooth muscle cells, suggesting its potential pharmacological relevance for the prevention or treatment of calcium-dependent vascular disorders.
Conclusion
The present study demonstrated that ZS 22-12 possesses significant vasorelaxant activity in isolated rat thoracic aorta through modulation of multiple calcium-regulating transport systems involved in vascular smooth muscle contraction. In vitro pharmacological experiments revealed that ZS 22-12 effectively inhibited both KCl-induced and phenylephrine-induced contractions in a concentration-dependent manner, indicating the involvement of voltage-dependent L-type Ca²⁺ channels as well as GPCR-associated receptor-operated calcium signaling pathways.Comparative analysis with verapamil and phentolamine further confirmed that the inhibitory effects of ZS 22-12 are closely associated with calcium influx and receptor-mediated intracellular calcium mobilization mechanisms. Molecular docking studies additionally demonstrated stable interactions of ZS 22-12 with several key calcium-regulating proteins, including Ca²⁺ L-type and R-type ion channels, SERCA, RyR, NCX, and Ca²⁺-ATPase. The obtained binding affinities and interaction profiles suggest that hydrophobic, hydrogen bonding, and electrostatic interactions contribute to stabilization of the ligand–protein complexes.
Acknowledgement
The author would like to thank Plantcytoprotectors Laboratory and Aripov Takhir Fatikhovich for their major recommendations.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
- Shakhnoza Bakirova: Conceptualization, Visualization.
- Izzatullo Abdullaev: Methodology, Data Analysis, Writing – Original Draft Preparation.
- Ergashev Shahzod.: Visualization.
- Shakhlo Ismailova: Visualization.
- Ulugbek Gayibov: Supervision, Writing – Review & Editing.
- Sirojiddin Omonturdiyev: Supervision.
- Kamila Eshbakova: Compound Synthesis.
- AllamuratovMirtaza: Funding Acquisition, Resources.
References
- Kapolou A, Karantonis HC, Rigopoulos N, et al. Association of mean daily polyphenols intake with Mediterranean diet adherence and anthropometric indices in healthy Greek adults: A retrospective study. Appl Sci. 2021;11:4664.
CrossRef - Li Q, Gao B, Siqin B, et al. Gut microbiota: A novel regulator of cardiovascular disease and key factor in the therapeutic effects of flavonoids. Front Pharmacol. 2021;12:651926.
CrossRef - Taguchi C, Kishimoto Y, Fukushima Y, et al. Dietary intake of total polyphenols and the risk of all-cause and specific-cause mortality in Japanese adults: The Takayama study. Eur J Nutr. 2020;59:1263–1271.
CrossRef - Zujko ME, Waskiewicz A, Witkowska AM, et al. Dietary total antioxidant capacity and dietary polyphenol intake and prevalence of metabolic syndrome in Polish adults: A nationwide study. Oxid Med Cell Longev. 2018;2018:7487816.
CrossRef - Mirsafaei L, Reiner Z, Shafabakhsh R, et al. Molecular and biological functions of quercetin as a natural solution for cardiovascular disease prevention and treatment. Plant Foods Hum Nutr. 2020;75:307–315.
CrossRef - Pechanova O, Dayar E, Cebova M. Therapeutic potential of polyphenols-loaded polymeric nanoparticles in cardiovascular system. Molecules. 2020;25:3322.
CrossRef - Jeon S, Lee S, Choi Y, et al. The effects of polyphenol-rich black elderberry on oxidative stress and hepatic cholesterol metabolism. Appl Sci. 2021;11:10018.
CrossRef - Makhmudov L, Tagayalieva N, Abdullaev I, et al. In silico pharmacokinetics and in vivo anti-inflammatory activity of C-4 composition. Biomed Pharmacol J. 2026;19.
CrossRef - Sirojiddin O, Izzatullo A, Ulugbek G, et al. Cardiovascular effects of Matricariachamomilla extract: Calcium channel modulation and vasorelaxant activity. Naturwissenschaften. 2026;113:14.
CrossRef - Abdurazakova I, Zaynabiddinov A, Abdullaev I, et al. Pharmacological evaluation of F45 on the cardiovascular system using in vitro, in vivo models and molecular dockings. Trends Sci. 2025;22:10924.
CrossRef - Gayibov UG, Gayibova SN, Abduazimova DS, et al. Antioxidant and cardioprotective properties of polyphenolic plant extract of Rhus glabra L. Plant Sci Today. 2024;11:2348–1900.
CrossRef - Sayidaliyeva R, Kadirova S, Zaynabiddinov A, et al. A-51 as a natural calcium channel blocker: An integrative study targeting hypertension. Trends Sci. 2025;22:10760.
CrossRef - Khasanov A, Abdullaev I, Kadirova S, et al. N-2 polyphenol targets vascular calcium channels to exert antihypertensive effects: In vitro and in vivo evaluation. Trends Sci. 2025;22:10782.
CrossRef - Sodiqova S, Kadirova S, Zaynabiddinov A, et al. Channelopathy activity of A-41 (propyl ester of gallic acid): Experimental and computational study of antihypertensive activity. Trends Sci. 2025;22:10496.
CrossRef - Abdullaev A, Abdullaev I, Bogbekov A, et al. Antioxidant potential of Rhodiolaheterodonta extract: Activation of Nrf2 pathway via integrative in vivo and in silico studies. Trends Sci. 2025;22(5):9521. doi:10.48048/tis.2025.9521.
CrossRef - Frąk W, Wojtasińska A, Lisińska W, et al. Pathophysiology of cardiovascular diseases: Molecular mechanisms of atherosclerosis, hypertension, and coronary artery disease. Biomedicines. 2022;10(8):1938. doi:10.3390/biomedicines10081938.
CrossRef - Wu X, Sha J, Yin Q, et al. Global burden of hypertensive heart disease. Sci Rep. 2025;15:14594. doi:10.1038/s41598-025-14594.
CrossRef - Teppo K, Airaksinen KEJ, Vasankari T, et al. Long forms of cardiac troponin T for myocardial infarction diagnosis. Eur Heart J. 2026;47(4):490–499. doi:10.1093/eurheartj/ehac490.
CrossRef - Abdullaev AA, Inamjanov DR, Abduazimova DS, et al. Sílybummarianum’s impact on physiological alterations and oxidative stress in diabetic rats. Biomed Pharmacol J. 2024;17(2):1291–1300.
CrossRef - Inomjonov D, Abdullaev I, Omonturdiev S, et al. In vitro and in vivo studies of Crataegus and Inula helenium extracts. Trends Sci. 2025;22(3):9158.
CrossRef - Khasanov A, Abdullaev I, Kadirova S, et al. Polyphenol N-2 targets vascular calcium channels. Trends Sci. 2025;22(12):10782.
CrossRef - Sodiqova S, Kadirova S, Zaynabiddinov A, et al. Channelopathy activity of A-41 (propyl ester of gallic acid): Experimental and computational study of antihypertensive activity. Trends Sci. 2025;22(9):10496.
CrossRef - Zoirovich OS, Zaynabiddinov AI, Ibragimov DR, et al. The effect of Ajugaturkestanica on rat aortic smooth muscle ion channels. Biomed Pharmacol J. 2024;17(2).
CrossRef - Zaripova M, Abdullaev I, Bogbekov A, et al. In vitro and in silico studies of Gnaphalium extract. Trends Sci. 2025;22(8):10098.
CrossRef - Gayibov UG, Gayibova SN, Abduazimova DS, et al. Antioxidant and cardioprotective properties of polyphenolic extract of Rhus glabra L. Plant Sci Today. 2024;11(3):2348–1900.
CrossRef - Aripov TF, Gayibov UG. Antiradical and antioxidant activity of the preparation “Rutan” from Rhuscoriaria L. J Theor Clin Med. 2023;4:164–170.
- Mahmudov AV, Abduraimov OS, Erdonov SB, et al. Bioecological features of Nigella sativa L. Plant Sci Today. 2022;9(2):421–426.
CrossRef - Mamajanov M, Abdullaev I, Sotimov G, et al. Mitochondrial and pharmacokinetic insights into pentahydroxyflavanone. Trends Sci. 2025;22(12):10984.
CrossRef - Aripov TF, Gayibov UG, Gaibova SN, et al. In vitro antioxidant and antiradical activity of total polyphenols of tannic sumach (Rhuscoriaria L.) leaves. KhimRastitelnogoSyr’ya. 2024;4:138–148.
CrossRef - Gayibov UG, Komilov EJ, Rakhimov RN, et al. Influence of a new polyphenol compound from Euphorbia plant on mitochondrial function. J MicrobiolBiotechnol Food Sci. 2019;8(4):1021–1025.
CrossRef - Gayibov U, Gayibova S, Mamurjon K, et al. Influence of quercetin and dihydroquercetin on rat liver mitochondria. J MicrobiolBiotechnol Food Sci. 2021;11(1):e2924.
CrossRef - Pozilov MK, Gayibov U, Asrarov MI, et al. Mitochondrial alterations under diabetes. J MicrobiolBiotechnol Food Sci. 2022;12(2):e2224.
CrossRef - Gilbert G, Demydenko K, Dries E, et al. Calcium signaling in cardiomyocyte function. Cold Spring HarbPerspect Biol. 2020;12(3):a035428.
CrossRef - Schneider T, Dibue-Adjei M, Neumaier F, et al. R-type voltage-gated Ca²⁺ channels in cardiac and neuronal rhythmogenesis. CurrMolPharmacol. 2015;8(1):102–108.
CrossRef - Ottolia M, John S, Hazan A, et al. The cardiac Na⁺–Ca²⁺ exchanger: From structure to function. Compr Physiol. 2021;12(1):2681–2717. doi:10.1002/cphy.c200031.
CrossRef - Pott C, Eckardt L, Goldhaber JI. Triple threat: The Na⁺/Ca²⁺ exchanger in the pathophysiology of cardiac arrhythmia, ischemia and heart failure. Curr Drug Targets. 2011;12(5):737–747.
CrossRef - Lanner JT, Georgiou DK, Joshi AD, et al. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring HarbPerspect Biol. 2010;2(11):a003996.
CrossRef - Laver DR. Ca²⁺ stores regulate ryanodine receptor Ca²⁺ release channels via luminal and cytosolic Ca²⁺ sites. Biophys J. 2007;92(10):3541–3555. doi:10.1529/biophysj.106.099028.
CrossRef - Magrone T, Magrone M, Russo MA, et al. Recent advances on the anti-inflammatory and antioxidant properties of red grape polyphenols: In vitro and in vivo studies. Antioxidants. 2019;9:35.
CrossRef - Mishra R, Jakhar M, Dhiman R, et al. The therapeutic potential of novel drug targets in metabolic disorders: A review. Biomed Pharmacol J. 2026;19(1).
CrossRef

