
Azam Asemi Rad1, Mohammad Hasan Heidari1, Abbas Aliaghaei1, Mehdi Eskandarian Broujeni3, Amir Shojaei4, Hojjat-Allah Abbaszadeh2, Fatemeh Shaerzadeh5 and Yousef Sadeghi2
1Anatomy and cell biology department, School of medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran.
2Hearing disorders research center, Shahid Beheshti University of Medical Sciences, Tehran, Iran.
3Stem cells and regenerative medicine, Faculty of medical biotechnology, National institute of genetic engineering and biotechnology, Tehran, Iran.
4Department of Physiology, Faculty of Medical Sciences, Tarbiat Modares University, Tehran, Iran.
5Molecular Medicine Research Center, Hormozgan University of Medical Sciences, Bandar Abbas, Iran; Department of Physiology, faculty of medicine, Hormozgan University of Medical Sciences, Bandar Abbas, Iran.
Corresponding Author E-mail: dr.ysadeghi@yahoo.com
DOI : https://dx.doi.org/10.13005/bpj/1146
Abstract
Nowadays, cell replacement using stem cells has been considered as an important potential therapy for treatment of Parkinson’s disease. Since the production of functional dopaminergic neurons is the first step in cell-replacement therapy, herein, we aimed to develop a high efficiency method to generate dopaminergic neurons from adipose derived stem cells (ADSCs) obtained from the adult adipose tissue. ADSCs were isolated and treated with growth factors for 2 weeks. Next, immunoreactivity for neuronal markers nestin, β-tubulin3, and microtubule-associated protein 2 and dopaminergic specific marker tyrosin hydroxylase was evaluated. Moreover, mRNA expression nestin, MAP2, PITX3, TH and KCNH5 was measured using quantitative real time PCR. Electrophysiological properties and ability of dopamine secretion were evaluated using current clamp and HPLC methods Our data. Revealed that ADSCs showed the morphological changes following neuronal induction after 14 weeks. Furthermore, ADSCs-derived dopaminergic neurons were immunopositive for neuronal and dopaminergic-specific markers. In addition, mRNA expression of nestin, MAP2, PITX3, TH and KCNH5 significantly increased in ADSCs-derived dopaminergic neurons. Finally, electrophysiological results confirmed that these neurons are able to excitation and they could produce action potential. Moreover, interestingly, HPLC analysis indicated that this neuronal cells have capacity of dopamine production and secretion that was in line with over-expression of TH enzyme in these cells. Although we provided a collection of data about the functional dopaminergic neurons derived from ADSCs, further in vivo studies are required to confirm the suitability of these dopaminergic cells for cell replacement in disorder related to dopaminergic neurons lesions.
Keywords
dopamine Parkinson disease; neural Stem cell; tyrosine hydroxylase;
Download this article as:| Copy the following to cite this article: Rad A. A, Heidari M. H, Aliaghaei A, Broujeni M. E, Shojaei A, Abbaszadeh H, Shaerzadeh F, Sadeghi Y. In Vitro Differentiation of Adipose Derived Stem Cells Into Functional Dopaminergic Neurons. Biomed Pharmacol J 2017;10(2). |
| Copy the following to cite this URL: Rad A. A, Heidari M. H, Aliaghaei A, Broujeni M. E, Shojaei A, Abbaszadeh H, Shaerzadeh F, Sadeghi Y. In Vitro Differentiation of Adipose Derived Stem Cells Into Functional Dopaminergic Neurons. Biomed Pharmacol J 2017;10(2). Available from: http://biomedpharmajournal.org/?p=15127 |
Introduction
Parkinson’s disease (PD) known as a neurodegenerative disorder whose symptoms of resting tremor, rigidity, and bradykinesia are caused by the degeneration of dopamine (DA) neurons in the midbrain. Although the symptoms of early-stage PD can be alleviated by a variety of palliative treatments, efficacy declines and treatment- related side effects emerge with the inexorable progression of the illness. Consequently, cell replacement using stem cells remains an important potential therapy, particularly at later stages of the disease.1-3 multi-potent differentiation and self- renew of Mesenchymal stem cells (MSCs) made these cells an appropriate source for the regeneration of nerve tissue.4 On the other hand, adipose tissue has been known as multi-potent stem cells assigned the generic nomenclature, adipose-derived stem cells (ADSCs).5,6 These cells can be differentiated to various mesenchymal tissues such as chondrocytes, myocytes ,adipocytes and osteoblasts.7-9 Differentiation of these cells into neural cells provide advantages for treatment of neurodegenerative diseases. To date, the ability of ADSCs differentiation towards non-mesenchymal lineages have not been proved. Recently, it has been reported that murine and human ADSCs can be induced to phenotypic as well as morphologic changes coherent with neuronal differentiation.10 One promising source of renewable cells for transplantation in PD are human neural progenitor cells (hNPs) derived from pluripotent human embryonic stem (hESCs) cell lines. Their success, however, greatly depends on the discovery of ways to promote their proper differentiation into midbrain dopamine (mDA) neurons. There are findings about derivation of DA neurons from ESCs.11-13 Even so, using of ESCs for transplantation can be led to immune rejection because of forming teratomas.
Neural stem cells (NSCs) are capable to differentiate into all types of brain cells.14 In spite of using NSCs for neural disorders therapy , these cells have some downsides regard to effectiveness and lineage polarization.2 MSCs are capable to generate DA neurons so they are useful for regenerating medicine.15-17 Several studies have shown that MSCs can transdifferentiate into the functional neural cells.18-20 Also several studies have been demonstrated that DA-type cells can be generated at low efficiency from MSCs in rodent models.21,22 In this study our aim was to stablish a high efficacy method to generate DA cells from ADSCs by using adult adipose tissue.
Materials and Methods
Isolation and Culture of Adscs
Adult male rats were sacrified by CO2 and their pretestis adipose tissue was removed. The samples were digested with 0.1% collagenase type I (Sigma—Aldrich, St. Louis, MO) by gently shaking at 37 °C for 30 min. For obtaining the stromal cell fraction, these samples centrifuged at 1500 rpm for 5 min . The pellet was filtered with 70 mm nylon mesh filter, and suspended in phosphate- buffered saline (PBS). The cell suspension was cultured at 37 °C in a control medium (Dulbecco’s modified Eagle medium: DMEM, 10% fetal bovine serum (FBS), 100 units/ml of penicillin, 100 mg/ml of streptomycin). After 48hrs, to remove the debris and red blood cells, the culture media was changed. ADSCs were cultured and expanded in culture medium, and used for the experiments at passages 1-3. For collecting the conditioned media of ADSC (ADSC-CM) for treatment, ADSC were cultured in serum free medium. After 48hrs, medium was collected, filtered and kept at -20°C for future uses.
Osteogenic Differentiation and Adipogenic Differentiation
For osteogenic differentiation, 3 × 103 cells/cm² were plated in DMEM supplemented with 10% FBS, 10 mM β-glycerophosphate, 60 μM ascorbic acid, and 10-7 M dexamethasone (Sigma Aldrich, Belgium), and cultured for 3 to 4 weeks, changing the medium every 3-4 days. To prove osteogenic differentiation, the cultures were fixed and stained with Alizarin red (Sigma Aldrich, Belgium).
The adipocyte differentiation performed by culturing 3 × 103 cells in DMEM/F12 supplemented by 10% FBS, 60 μM indomethacin, 10-6 M dexamethasone and 5 μg/ml insulin and 0.5 mM isobutylmethylxanthine(IBMX) (Sigma). Then the cells were cultured, displacing the medium every 3–4 days. After 3 to 4 weeks, cells were consist of neutral lipids in fat vacuoles; they were fixed by using 10% formalin and stained with fresh oil red-O solution (Sigma Aldrich, Belgium).
Dopaminergic Differentiation
In our study, we induced ADSCs into dopaminergic neurons by two steps. Firstly, ADSCs were induced to become neuronal cells as well as neural stem cells. ADSCs were pre-induced by overnight treatment with bFGF (20 ng/ml), butylated hydroxyanisole (BHA, 200 μM) and B27 (1%) in DMEM/F12 and 20% fetal bovine serum (FBS). In the second step, dopaminergic differentiation was done by incubation of differentiated neural cells in dopaminergic culture medium containing Shh (100 ng/ml), FGF8 (100 ng/ml) and B27 (1%) in DMEM/F12 and 5% FBS for 14 days.
Patch Clamp Recording in Cell Cultures
Cells were prepared for electrophysiological recording at two weeks after dopaminergic differentiation. Neuronal-like cells were seeded on poly D-lysine coated coverslips. Cells were incubated at room temperature (23–25°C) for 30 minutes and then transferred to a submerged recording chamber that continually perfused at 1.5–2.5 ml/min with standard artificial cerebrospinal fluid (ACSF). ACSF was bubbled with 95% O2–5% CO2 continuously and contained 125 NaCl, 3 KCl, 1.25 NaH2PO4, 25 NaHCO3, 10 D-Glucose, 2 CaCl2 and 1.3 MgCl2. The recording chamber was rised on a fixed-stage upright microscope (Axioskop 2 FS MOT; Carl Zeiss, Göttingen, Germany). An IR-CCD camera (IR-1000, MTI, USA) with a 40× water immersion objective lens was used for visualization of cells. Patch pipettes were pulled from borosilicate glass (1.5-mm outer diameter; Harvard Apparatus, Edenbridge, UK) using a horizontal puller (P-97, Sutter Instrument, Novato, CA, USA) and filled with an intracellular solution that contained (in mM) 115 K-gluconate, 20 KCl, 10 HEPES, 2 EGTA, 10 disodium-phosphocreatine, 2 MgATP and 0.3 NaGTP. pH was adjusted to 7.25–7.30 and osmolality to 285–290 mOsm. Electrode tip resistance in the bath was typically 6–8 MΩ and series resistance ranged from 10–30 MΩ. Data were low-pass filtered at 10 kHz and acquired at 10 kHz with a Multiclamp 700B amplifier equipped with a Digidata 1440 A/D converter (Molecular Devices, Sunnyvale, CA, USA). The signal was recorded on a PC using the Axon pClamp 10 acquisition software (Molecular Devices, Palo Alto, CA).
Immunocytochemistry
Cells were washed with phosphate buffered saline (PBS) and then fixed by paraformaldehyde (PFA, 4%) for immunofluorescence, followed by washing with PBS (3 times) and air drying. Then slides were blocked with normal blocking serum (%10) in PBS for 40 min and washed by PBS, and incubated by primary antibody including anti TH (ABCAM), anti Nestin (ABCAM) and anti β-tubulin3 (ABCAM) in 1.5% normal blocking serum in PBS overnight. The slide was washed 3 times with PBS and then incubated with fluorescein isothiocyanate (FITC) or TRITC-conjugated secondary antibody (ABCAM) for 60 min. Fluorescence microscopy was used for visualizing immunoreactive cells.
RNA Extraction and Reverse Transcriptase Polymerase Chain Reaction
Total RNA was extracted using High Pure RNA Isolation kit, according to the manufacturer’s instructions (Roche, Basel, Switzerland) followed by transcription of 1 ug of total RNA to cDNA using murine leukemia virus (MuLV) reverse transcriptase (Fermentas, Lithuania) in the presence of random hexamers and RNase inhibitor. Afterwards, obtained cDNA was boosted by 35 cycles of denaturation at 94°C for 30 second, annealing at 60°C for 30 second and extension at 72°C for 30 s using specific primers (Table 1). Size-fractionating of PCR products performed by 2% agarose gel electrophoresis.
Quantitative Real-Time PCR
For quantitative real-time PCR (qPCR) analysis, specific primers for Nestin, MAP2, PITX3, KCNH5 and TH genes and actin as internal control were used (Table 1). Reactions were accomplished using SYBR® Premix Ex Taq™ II (TAKARA BIO INC.) on a Rotor-GeneTM 6000 real-time PCR machine (Corbett Research, Qiagen, Germany). Initial denaturation was performed at 95ºC for 15 minutes. Then 45 cycles of denaturation at 95 ºC for 10 seconds was used under primer specific conditions (Table 1) as well as extension at 72ºC for 20 seconds.
Table 1: Total cellular RNA was analyzed using the primers against several neural markers.
| Primer name | sequence | Size(bp) | annealing | accesslon | |
| Rat actin | F | CCTGGGCATGGAGTCCTGT | 153 | 61OC × 20 sec | NM_OO1127449.1 |
| R | ATCTCCTTCTGCATCCTGTCG | ||||
| Rat PITX3 | F | AGCACAGCGACTCAGAAAAGG | 225 | 61OC × 20 sec | NM_019247.1 |
| R | TCTTGAACCACACCCGCACG | ||||
| Rat TH | F | AGCTCCTGCACTCCCTGTCA | 152 | 61OC × 20 sec | NM_012740.3 |
| R | GGCATAGTTCCTGSGCTTGTC | ||||
| Rat Nestin | F | CCTCAAGATGTCCCTTAGTCTG | 114 | 61OC × 20 sec | NM_001308239.1 |
| R | TCCAGAAAGCCAAGAGAAGC | ||||
| Rat MAP2 | F | CAAACGTCATTACTTTACAACTTGA | 122 | 61OC × 20 sec | NM_013066.1 |
| R | CAGCTGCCTCTGTGAGTGAG | ||||
| Rat KCNH5 | F | GATCCTGGCCGCTGCTCTCC | 101 | 57 OC × 20 sec | NM_133610.2 |
| R | ACTGGAGCGCCTGACGATGT | ||||
High-Performance Liquid Chromatography Analysis of Dopamine Concentration
For dopamine measurement High performance liquid chromatography-electrochemical detector (HPLC-ECD) was used. Briefly, under dopaminergic differentiation ADSCs were washed in cold PBS prior to depolarization with 50 mM KCL in Hanks’ balanced salt solution (HBSS) for 15 minutes. Subsequently, samples were collected, sonicated and centrifuged for 20 min at 15000 rpm at 4°C. Then the supernatant was collected as well as transferred to a clean black tube. After all 50 microliters of 0.1 M perchloric acid (PCA; Merck) was added. Next, 250 μl volume of sample was transferred to a 1.5 ml polypropylene tube and 10 μL methanol was added. After word the sample was vortex-mixed for 30 s and was centrifuged for 2 min at 12000 rpm. A volume of 10 μl of supernatant was injected into the chromatograph. A LKB Bromma 2150 solvent delivery pump (LKB, Sweden), an EICOM ECD-100 electrochemical detector at +600 mV with highest sensitivity, and Clarity Lite HPLC software (Data Apex, Petrzilkova, Czech Republic) were utilized. Knauer loop injector was used for applying samples with an efficient volume of 100 μl. A C8e Monolithic column (Chromolith, 100 × 4.6 mm) protected by a Chromolith C18e (5 × 4.6 mm) was used for the chromatographic separation (Merck, Germany). The mobile phase consist of 500 ml of 50 mM sodium dihydrogen phosphate, 25 ml of acetonitrile, 5 mg of SLS and 5 mg EDTA, was adjusted at pH 3.1 with concentrated phosphoric acid. Analysis were run at flow rate of 1.5 ml/min at ambient temperature. A direct comparison to the dopamine peak area of the concentration of 160 ng/ml (i.e. 12000) was used to quantify dopamine concentration.
Data Analysis
The data analysis was represented as the mean ± SEM. ANOVA was used to comparison between groups as well as Tukey’s multiple comparison test (specific post-hoc test). The comparative quantitation was performed between selected groups by using REST 2009 (Relative Expression Software Tool, Qiagen) based on Pair Wise Fixed Reallocation Randomization Test (Pfaffl et al., 2002). The statistical significances were achieved when P<0.05.
Results
Characterization of Adscs
In the primary culture, ADSCs isolated from rat pretestis adipose tissue, appeared as a fairly uniform population that displayed a fibroblast-like morphology and they well expanded in vitro (Fig.1A). Immunophenotyping of ADSCs displayed these cells as positive cells for mesenchymal stem cells markers such as CD105 and CD73. At the same time, ADSCs were negative for the hematopoietic surface marker CD45 (Fig.1B).
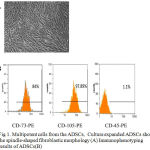 |
Figure 1: Multipotent cells from the ADSCs, Culture expanded ADSCs show the spindle-shaped fibroblastic morphology (A) Immunophenotyping results of ADSCs(B)
|
Mesodermal Lineage Differentiation of Adscs
ADSCs did not spontaneously differentiate during in vitro culture expansion. During in vitro differentiation of ADSCs with osteogenic medium, calcified ECM deposition was detected by Alizarin Red staining, as shown in Fig.2A. Furthermore, using Oil Red O staining exhibited intracellular accumulation of lipid droplets that supported adipogenic differentiation (2qFig.2B).
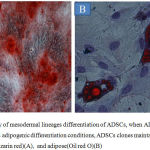 |
Figure 2: Capacity of mesodermal lineages differentiation of ADSCs, when ADSCs exposed to osteogenic and adipogenic differentiation conditions, ADSCs clones maintain osteogenic(Alizarin red)(A), and adipose(Oil red O)(B)
|
Morphological Changes Followed by Neuronal Induction of Adscs
Following neuronal pre induction by FGF-2 (20ng/ml) and B27, ADSCs displayed rapid changes in cellular morphology. Following neuronal pre induction for 24 h, the cytoplasm of ADSCs retracted towards the nucleus and forming condensed cell bodies with cytoplasmic extensions. The cell bodies increasingly converted to spherical and refractile shape, showing a neuronal perikaryal appearance. Although the onset of these changes varied slightly between experiments, we saw dramatic rearrangements in cytoskeletal and membrane appearance in ADSCs cultures within 24 h following neuronal pre induction (Fig. 3A and 3B). In the next step, we induced dopaminergic differentiation by incubation of ADSCs-derived neuron like cells in dopaminergic culture medium containing FGF8 (100ng/ml) and Shh (100ng/ml). As depicted in Fig. 3B, following the addition of inducing agents into the medium, morphology of cells deeply changed. Multiple neuritis were extended from the cell body alongside a single long axon-like processes in some differentiated cells. After 5-7 days of dopaminergic differentiation, some colonies with neurospheres morphology similar to the NSCs colonies were observed (Fig.4A). Furthermore, to determine if the cultured spheres derived from ADSCs resembled to neurospheres, we evaluated the NSCs-specific antigen nestin in ADSCs colonies.23 As shown in Fig. 4B, 4C and 4D, immunocytochemistry data revealed that most of the cells within a single sphere express nestin.
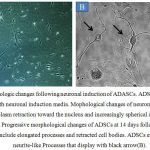 |
Figure 3: Morphologic changes following neuronal induction of ADASCs. ADSCs after 24 h of culture with neuronal induction media.
|
Mophological changes of neuronal differentiation include cytoplasm retraction toward the nucleus and increasingly spherical appearance of cell bodies(A). Progressive morphological changes of ADSCs at 14 days following neuronal induction include elongated processes and retracted cell bodies. ADSCs extend long thin neurite-like Processes that display with black arrow(B).
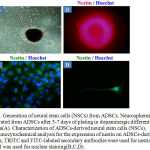 |
Figure 4: Generation of neural stem cells (NSCs) from ADSCs. Neurospheres generated from ADSCs after 5-7 days of plating in dopaminergic differentiation media(A).
|
Characterization of ADSCs-derived neural stem cells (NSCs), Immunocytochemical analysis for the expression of nestin on ADSCs-derived NSCs, TRITC and FITC-labeled secondary antibodies were used for nestin. DAPI was used for nuclear staining(B,C,D).
Adscs-Derived Dopaminergic Neurons Immunopositivity for Neuronal and DA-Specific Markers
Fourteen days after dopaminergic differentiation, immunocytochemical staining analysis showed that ADSCs-derived dopaminergic neurons were positive for neuronal markers β-tubulin3 and MAP2 (Fig.5A, 5B). ADSCs-derived dopaminergic neurons were also immunopositive for catecholaminergic rate limiting synthesizing enzyme TH as shown in Fig.6A. Moreover, double immunocytochemical staining showed that ADSCs-derived dopaminergic neurons were positive for neuronal marker NeuN and dopaminergic marker TH as it can be seen in Fig.6B.
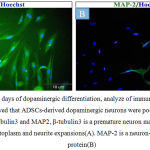 |
Figure 5: After 14 days of dopaminergic differentiation, analyze of immunocytochemical staining showed that ADSCs-derived dopaminergic neurons were positive for neuronal markers β-tubulin3 and MAP2, β-tubulin3 is a premature neuron marker and strongly expressed in cytoplasm and neurite expansions(A). MAP-2 is a neuron-specific cytoskeletal protein(B)
|
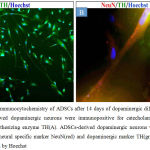 |
Figure 6: Immunocytochemistry of ADSCs after 14 days of dopaminergic differentiation. ADSCs-derived dopaminergic neurons were immunopositive for catecholaminergic rate limiting synthesizing enzyme TH(A). ADSCs-derived dopaminergic neurons were double stained for neural specific marker NeuN(red) and dopaminergic marker TH(green). Nuclei were labeled by Hoechst
|
Adscs-Derived Dopaminergic Neurons Expressed Neuronal and DA-Specific Genes
Based on RT-PCR results, differentiated ADSCs expressed neuronal and dopaminergic markers Nestin, MAP2, KCNH5, PITX3 and TH (Fig.7). These observations were also confirmed using qPCR method. Based on the results, mRNA levels of Nestin and MAP2 increased 5.3 (P<0.05) and 14.42 (P<0.01) time compared to the non-differentiated cells, respectively (Fig. 7A and 7B). Furthermore, measurement of mRNA level of the ether-a`-go-go potassium channel (KCNH5) demonstrated that increasing the expression of this gene in differentiated cells compared to control group was significant statistically (3.68 fold compared to control cells (P<0.05) (Fig.7C)). Moreover, the expression levels of dopaminergic markers PITX3 and TH were up regulated 13.67 (P<0.01) and 7.01 (P<0.05) times, respectively, in comparison with non-differentiated cells (Fig.7D and 7E).
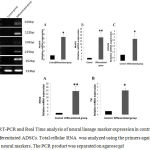 |
Figure 7: RT-PCR and Real Time analysis of neural lineage marker expression in control ADSCs and differentiated ADSCs. Total cellular RNA was analyzed using the primers against several neural markers. The PCR product was separated on agarose gel
|
Excitable Properties of Adscs-Derived Dopaminergic Neurons
After two weeks of transfection, functional assessment of induced neurons was done by patch clamp recording. We used 10 neuron-like cells to evaluate resting membrane potential and the capability to generate action potential. The current clamp mode was applied to examine shifts in membrane voltage in response to injection of depolarizing currents. Before injection of depolarizing currents, voltage was adjusted to -60 mV and then current steps ranging between 50 and 200 pA (600 ms) were applied in 50 pA increments to record membrane potential. We observed a single action potential-like event following current injection in the induced neurons with maximum amplitude of about 10 mV , implying the induced neurons express functional voltage-gated ion channels (Fig.8).
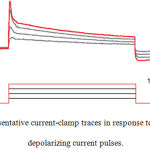 |
Figure 8: Representative current-clamp traces in response to injection of depolarizing current pulses.
|
Dopamine Secreted from Adscs-Derived Dopaminergic Neurons
For examining the capability of ADSCs-derived dopaminergic neurons to release dopamine Reverse-phase HPLC was used. In response to a K+ depolarizing stimulus, dopamine (16.2 ng/ml) was released into the medium After two weeks of differentiation. This indicated that functional dopaminergic neurons with the ability of dopamine secretion were present in the culture (Fig.9).
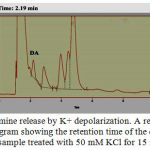 |
Figure 9: Dopamine release by K+ depolarization. A representative chromatogram showing the retention time of the dopamine peak a sample treated with 50 mM KCl for 15 minutes
|
Discussion
ADSCs transplantation have attracted attention as potentiality therapy for a wide variety of diseases including diabetes, trauma and neurodegenerative disorders.24,25 In this regard, as the second most common progressive neurodegenerative disorder, todays, many attempts have been made to recreate the nigrostriatal pathway by transplantation of dopaminergic neurons for treatment of PD. So, the production of functional dopaminergic neurons is the first step in cell-replacement therapy in this dopaminergic neuron lesion-related neurodegenerative disorder. Accordingly, in this study following the efforts that have been made in this field, we induced a functional dopaminergic neurons with the potency of DA secretion alongside excitability properties.
The identification of the active neurogenesis capacity in specific areas of adult brain opens up a new window in neurological and neurodegenerative disorders research to look at the possible replacement therapies. However, one potential issue ahead in this context was the finding of feasible population cells for replacement therapies and transplantation in specific regions of the brain that involve in neurodegeneration. Accordingly, it seems that using ADSCs are a suitable source of cells for transplantation as they are available and could be simply isolated beside their ability for being rapidly expanded and differentiated into neuronal cells.4,5,26 As the data revealed a 14-day period of incubation with growth factors led to induction of functional stem cell-derived dopaminergic neurons. To confirm this, we firstly evaluated specific neuronal markers tubulin3 and MAP2 in neurons derived from ADSCs. The MAP2 is a neuron-specific marker that stabilizes microtubules in the expanded dentrites of postmitotic neurons and is known as a neuronal differentiation protein.27 Moreover, as a cytoskeleton protein, tubulin3 is exclusively regarded as a neuronal marker.28 The results showed that induced dopaminergic neurons significantly expressed these two neuronal markers confirming their differentiation into the neuronal cells.
Secondly, we immunocytochemcally examined the dopaminergic neuron marker TH, a catecholaminergic rate limiting synthesizing enzyme and then, expression level of mRNAs of nestin, MAP2, KCNH5, PITX3 and TH were evaluated. As previously shown, mRNA levels of these neuronal markers significantly increased in neuronal cells derived from ADSCs. The KCNH5 is invovlved in setting the negative resting potentials which is necessary for neuronal survival and excitability.29 As it was shown, mRNA level of these potassium channels notably increased in the induced dopaminergic neurons indicating the ability of these cells for producing negative resting potential across the membrane as confirmed by electrophysiological test. Moreover, mRNA level evaluation of PITX3 revealed that expression of this factor significantly elevated in ADSCs-derived neurons verifying that they are dopaminergic. The PITX3 constitutively expressed in the substantianigra after birth and is involved in the long-term survival and maintenance of the midbrain DA neurons.30
Finally, result of the current clamp study confirmed that the neurons derived from ADSCs are able to excite and could produce action potential. In addition to excitation property, interestingly, HPLC analysis indicated that these neuronal cells have capacity of dopamine production and secretion that was in line with over-expression of TH enzyme in these cells. TH is known as the gold standard marker for identification of dopaminergic neurons catalyzing the transformation of L-tyrosine to L-3,4-dihydroxyphenylalanine.31 In addition to the capacity of DA production based on the expression of TH, the HPLC data confirmed that they could secrete DA to the extracellular space.
In conclusion, we provided a collection of data about the functional dopaminergic neurons derived from ADSCs. The results indicated that the neuronal cells could produce action potential and have some channels involving in negative resting potentials. Moreover, it was revealed that the differentiated neurons produced and secreted DA which could introduce them as a potential candidate for transplantation for cell therapy in PD. However, further in vivo studies are required to confirm the suitability of these ADSCs-derived dopaminergic cells for cell replacement in disorders related to dopaminergic neurons lesions.
Acknowledgements
This work was supported by hearing disorders research center, Shahid Beheshti University of Medical Sciences and this project is part of PhD student thesis of A. Asemi Rad
References
- Lindvall O., Björklund A. Cell therapy in Parkinson’s disease. NeuroRx. 2004;1(4):382-93.
- Snyder B. J., Olanow C. W. Stem cell treatment for Parkinson’s disease an update for 2005. Current opinion in neurology. 2005;18(4):376-85.
- Kondo T., Johnson S. A., Yoder M. C., Romand R., Hashino E. Sonic hedgehog and retinoic acid synergistically promote sensory fate specification from bone marrow-derived pluripotent stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(13):4789-94.
- Pittenger M. F., Mackay A. M., Beck S. C., Jaiswal R. K., Douglas R., Mosca J. D., et al. Multilineage potential of adult human mesenchymal stem cells. science. 1999;284(5411):143-7.
- Gimble J. M., Guilak F. Adipose-derived adult stem cells: isolation, characterization and differentiation potential. Cytotherapy. 2003;5(5):362-9.
- Strem B. M., Hicok K. C., Zhu M., Wulur I., Alfonso Z., Schreiber R. E., et al. Multipotential differentiation of adipose tissue-derived stem cells. The Keio journal of medicine. 2005;54(3):132-41.
- Zuk P. A., Zhu M., Mizuno H., Huang J., Futrell J. W., Katz A. J., et al. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue engineering. 2001;7(2):211-28.
- Halvorsen Y. D. C., Bond A., Sen A., Franklin D. M., Lea-Currie Y. R., Sujkowski D., et al. Thiazolidinediones and glucocorticoids synergistically induce differentiation of human adipose tissue stromal cells: biochemical, cellular and molecular analysis. Metabolism. 2001;50(4):407-13.
- Erickson G. R., Gimble J. M., Franklin D. M., Rice H. E., Awad H., Guilak F. Chondrogenic potential of adipose tissue-derived stromal cells in vitro and in vivo. Biochemical and biophysical research communications. 2002;290(2):763-9.
- Safford K. M., Hicok K. C., Safford S. D., Halvorsen Y. D. C., Wilkison W. O., Gimble J. M., et al. Neurogenic differentiation of murine and human adipose-derived stromal cells. Biochemical and biophysical research communications. 2002;294(2):371-9.
- Ben‐Hur T., Idelson M., Khaner H., Pera M., Reinhartz E., Itzik A., et al. Transplantation of human embryonic stem cell–derived neural progenitors improves behavioral deficit in parkinsonian rats. Stem cells. 2004;22(7):1246-55.
- Buytaert‐Hoefen K. A., Alvarez E., Freed C. R. Generation of tyrosine hydroxylase positive neurons from human embryonic stem cells after coculture with cellular substrates and exposure to GDNF. Stem cells. 2004;22(5):669-74.
- Iacovitti L., Donaldson A. E., Marshall C. E., Suon S., Yang M. A protocol for the differentiation of human embryonic stem cells into dopaminergic neurons using only chemically defined human additives: studies in vitro and in vivo. Brain research. 2007;1127:19-25.
- Lie D. C., Dziewczapolski G., Willhoite A. R., Kaspar B. K., Shults C. W., Gage F. H. The adult substantia nigra contains progenitor cells with neurogenic potential. The Journal of neuroscience. 2002;22(15):6639-49.
- Alhadlaq A., Mao J. J. Mesenchymal stem cells: isolation and therapeutics. Stem cells and development. 2004;13(4):436-48.
- Barry F. P. Biology and clinical applications of mesenchymal stem cells. Birth Defects Research Part C: Embryo Today: Reviews. 2003;69(3):250-6.
- Caplan A. I., Bruder S. P. Mesenchymal stem cells: building blocks for molecular medicine in the 21st century. Trends in molecular medicine. 2001;7(6):259-64.
- Cho K. J., Trzaska K. A., Greco S. J., McArdle J., Wang F. S., Ye J. H., et al. Neurons derived from human mesenchymal stem cells show synaptic transmission and can be induced to produce the neurotransmitter substance P by interleukin‐1α. Stem cells. 2005;23(3):383-91.
- De Hemptinne I., Vermeiren C., Maloteaux J. M., Hermans E. Induction of glial glutamate transporters in adult mesenchymal stem cells. Journal of neurochemistry. 2004;91(1):155-66.
- Jin K., Mao X. O., Batteur S., Sun Y., Greenberg D. A. Induction of neuronal markers in bone marrow cells: differential effects of growth factors and patterns of intracellular expression. Experimental neurology. 2003;184(1):78-89.
- Fu Y. S., Cheng Y. C., Lin M. Y. A., Cheng H., Chu P. M., Chou S. C., et al. Conversion of human umbilical cord mesenchymal stem cells in Wharton’s jelly to dopaminergic neurons in vitro: potential therapeutic application for Parkinsonism. Stem cells. 2006;24(1):115-24.
- Guo L., Yin F., Meng H., Ling L., Hu-He T., Li P., et al. Differentiation of mesenchymal stem cells into dopaminergic neuron-like cells in vitro. Biomed Environ Sci. 2005;18(1):36-42.
- Fu L., Zhu L., Huang Y., Lee T. D., Forman S. J., Shih C. C. Derivation of neural stem cells from mesenchymal stem cells: evidence for a bipotential stem cell population. Stem cells and development. 2008;17(6):1109-22.
- Rammal H., Harmouch C., Lataillade J. J., Laurent-Maquin D., Labrude P., Menu P., et al. Stem cells: a promising source for vascular regenerative medicine. Stem cells and development. 2014;23(24):2931-49.
- Mariani E., Facchini A. Clinical applications and biosafety of human adult mesenchymal stem cells. Current pharmaceutical design. 2012;18(13):1821-45.
- Stewart M. C., Stewart A. A. Mesenchymal stem cells: characteristics, sources, and mechanisms of action. Veterinary Clinics of North America: Equine Practice. 2011;27(2):243-61.
- Soltani M. H., Pichardo R., Song Z., Sangha N., Camacho F., Satyamoorthy K., et al. Microtubule-associated protein 2, a marker of neuronal differentiation, induces mitotic defects, inhibits growth of melanoma cells and predicts metastatic potential of cutaneous melanoma. The American journal of pathology. 2005;166(6):1841-50.
- Katsetos C. D., Legido A., Perentes E., Mörk S. J. Class III β-tubulin isotype a key cytoskeletal protein at the crossroads of developmental neurobiology and tumor neuropathology. Journal of Child Neurology. 2003;18(12):851-66.
- Judge S. I., Smith P. J., Stewart P. E., Bever J., Christopher T. Potassium channel blockers and openers as CNS neurologic therapeutic agents. Recent patents on CNS drug discovery. 2007;2(3):200-28.
- Li J., Dani J. A., Le W. The role of transcription factor Pitx3 in dopamine neuron development and Parkinson’s disease. Current topics in medicinal chemistry. 2009;9(10):855-9.
- Goldstein D. S., Eisenhofer G., Kopin I. J. Sources and significance of plasma levels of catechols and their metabolites in humans. Journal of Pharmacology and Experimental Therapeutics. 2003;305(3):800-11.

