
R. Saxena1, R. Stephan2, S. K.Mishra4, S. Shukla3, D. P.Saxena1, S. O. Pratap4 and D. P.Singh4
1Vinayaka Mission University, Salem, T.N., India.
2Govt. Arts College, Department of Botany, Ariyaloor, T.N, India.
3Integral University, Lucknow, U.P., India.
4Central Avian Research Institute, Izatnagar, Bareilly, U.P., India.
Abstract
Immunocompetence is known to be controlled by genetics depending on the B-haplotype Major Histocompatibility complex (MHC), for instance and the selection for production traits. Genetic resistance to disease involves many facets of the body’s defense system and interactions, and it is extremely complex. Production traits had negative correlation to disease resistant traits. The aim of the current study was to estimate the immune responses against SRBC in selected lines, estimating the genetic trend and genetic correlation between male and female immune responses against SRBC. Humoral response was estimated in the form of titer against sheep RBC, each birds was received an intravenous injection of 0.1 ml of 0.5% suspension of packed SRBC in normal saline at 6 week of age Five day later the blood was collected from Jugular vein of each birds in the individual test tube and sera were collected. In the present investigation, chicks belonging to the selected lines of native chickens were used for the study of immunological traits viz., response to sheep RBCs, IgG concentrations. Finally, 1% sheep RBC suspension was prepared by mixing 1 ml of packed sheep RBCs and 99 ml of PBS which was then used for injection in the birds. The Average means for HA was observed (7.74±0.109) and IgG (1.94±0.72) in native breed. Sex wise HA results showed that male have higher mean (7.925 ± 0.1463) than females (7.57± 0.161) for HA while the same was also estimated for IgG by having mean (2.1194±8.9) and female (1.77±8.18). Test of Homogeneity of Variances was recorded as (0.39) for HA which was significant and IgG level (0.937).
Keywords
Immune competence; native breed; HA and HI titer; IgG
Download this article as:| Copy the following to cite this article: Saxena R, Stephan R, Mishra S. K, Shukla S, Saxena D. P, Pratap S. O. Assessment of Immunocompetence Status of Native Breed of Chickens. Biomed Pharmacol J 2012;5(2) |
| Copy the following to cite this URL: Saxena R, Stephan R, Mishra S. K, Shukla S, Saxena D. P, Pratap S. O. Assessment of Immunocompetence Status of Native Breed of Chickens. Biomed Pharmacol J 2012;5(2). Available from: http://biomedpharmajournal.org/?p=2508 |
Introduction
Increasing population of India with increased demand of nutritious food and shrinkage of land has drawn the attention of people towards the livestock production and poultry farming. Due to high nutritive value and less input cost, the poultry farming has grown up nicely. Hence, economics of poultry production can be effectively improved by decreasing the input costs. It has been amply proved that immunocompetence or resistance to infections/ diseases is under the control of gene or group of genes called gene complexes includes MHC, Interleukin, Interferon.
Immuno-competence is known to be controlled by genetics-depending on the B-haplotype (major Histocompatibility complex [MHC]), for instance and the selection for production traits. It has been shown that the proportion of immune cell subsets can be correlated to susceptibility and resistance to disease1,2.
Genetic enhancement of immune-competence without challenging animals with antigens may be a potential approach for improving health and production parameters as well as general disease resistance. Selection of chickens for serum IgG isotypes, which represents the response to a wide variety of unknown antigens, may be a new breeding strategy to improve resistance to infectious diseases. In this regard, genetic control of humoral and cell-mediated immunities and phagocytic ability of chicken lines selected for serum IgM and IgG has not been well investigated.
Avian immune system is a highly evolved and complex system involving different cell types and soluble factors that act in concert to elicit immune response against invading pathogens and thus play a central role in determining survivability. Immune response and disease resistance in chicken have been found to be affected by multiple genetic and environmental factors3. The latest discoveries in the area of molecular and immunegenetics have tremendously added to the vast knowledge of genome organizations, number of genes, their structures, expression and regulation and nucleotide polymorphism.
The major part of the immunological defense may be grouped in to three different and interdependent functional facets viz. cellular system, which provides cell mediated immunity or direct cellular intervention against pathogens, humoral system, which acts by producing humoral immunity or by antibody production, the non-specific system (e.g. complement system, phagocytosis etc.).
It has been demonstrated that different immune challenges produce diverse responses and so require different cells to be activated 4,5,6,7. For this reason, an assay intended to understand the current status of commercial poultry and the response capability of its immune system must comprise parameters that refer to these different pathways.
The majority of B-cells express major histocompatability (MHC) II molecules, and they are classified as antigen presenting cells. Early events in B-cell activation include antigen contact with specific B-cells, that leads to transmembrane signaling, increased expression of Class-II MHC molecules as secreted molecules in plasma and other bodily fluids, they are able to bind foreign antigens, either neutralizing them directly or indirectly through activation of the complement system or antibody dependent cell cytolysis by monocytic phagocytes8.
A meta-analysis of experiments increasing/decreasing clutch/brood size to alter parental effort found a significant negative relationship between parental effort and immune responses9.
Genetic resistance to disease involves many facets of the body’s defense system and interactions, and it is extremely complex. Production traits had negative correlation to disease resistant traits. In addition, disease resistant traits had low heritability, which made it difficult to use conventional breeding to improve the disease resistant traits. New opportunities to improve our understanding of the genetic nature of disease resistance have been produced through recent advances in molecular biology, gene mapping, and immunology which make selection for disease resistance possible in the future. However, testing and selection for disease resistance and improved immune responsiveness require knowledge of the genetic correlations.
This implies a cost of rather short duration, at least in physiologically non exhausted individuals; that is, when the strenuous activities cease, immune responses should be restored within days or weeks10.
Many factors regulating the immune responses in birds have been identified and characterized in last few decades. Macrophages are one of the most important immune-competent cells of the birds immune system, which are involved in initiating and directing the innate specific immune response. Selection based on these immune effectors may helps in enhancing disease resistance immunocompetence of the poultry flock. Cytokines play an important role in immune response. These are secreted by a variety of cells and two principal producers are the dendritic cells and macrophages. Cytokines are involved in development of cellular and humoral immune responses, induction of the inflammatory response, regulation of haematopoiesis, control of cellular proliferation and differentiation and healing of wounds. The present experiment studied the effect that selecting chickens divergently for antibody response to a single i.e. inoculation of SRBCs.
The genetics of a bird or flock has a profound impact on its ability to resist disease, because genetics define the maximum achievable performance level. Careful attention should be paid to genetics as an important component of a comprehensive disease management program including high-level biosecurity, sanitation, and appropriate vaccination programs. Some specific genes (e.g. MHC) are known to play a role in disease resistance, but resistance is generally a polygenic phenomenon. Future research directions will expand knowledge of the impact of genetics on disease resistance by identifying non-MHC genetic control of resistance and by further elucidating mechanisms regulating expression of genes related to immune response.
The immune system of an individual consists of three major facets antibody response (humoral immunity), T cell-mediated immunity and phagocytosis. Coordination of these systems enables an individual to resist infection and disease. Genetic control of these facets of the immune system may be independent of each other11, 12, 13, 14. The kinetics of antibody responses depends on the Ig isotype. The immune system of an individual consists of three major facets antibody response (humoral immunity), T cell-mediated immunity and phagocytosis. Coordination of these systems enables an individual to resist infection and disease. The challenging of these lines and the crosses between them with a range of challenging agents. The defense mechanisms were not only resource expensive, but depending on the genetic mechanisms involved, the populations were ranked depending on the challenging agent. Concomitant with the direct response of SRBC antibody titers to selection are correlated responses in production and disease-related traits15.
The level of maternal antibodies increases during the first 2 to 4 d of age16. Where after, it decreases until 2 to 4 wk of age17,18.
Genetic control of antibody response and cell mediated immunity is polygenic. The three major parts or systems of the immunological defense are under independent genetic control19. They may be distinguished as three sets of host response genes such as:
(a) Genes which control the susceptibility or resistance of an innate immunity.
(b) Genes which govern the specificity of acquired or adaptive immune response.
(c) Genes which affect the quality of specific immune response.
The control of immunosuppressive diseases is of prime importance for the nascent poultry industry in developing countries.
The aim of the current study was to estimate the immune responses against SRBC in selected lines, estimating the genetic trend and genetic correlation between male and female immune responses against SRBC.
Estimation of Humoral Immune Responses
Humoral response was estimated in the form of titer against sheep RBC, each birds was received an intravenous injection of 0.1 ml of 0.5% suspension of packed SRBC in normal saline at 6 week of age Five day later the blood was collected from Jugular vein of each birds in the individual test tube and sera were collected. The total antibody titer was determined by haemagglutination test perfumed in micro titer plate from individual serum sample two fold serial dilutions was carried out in normal saline solution and equal amount of 1% SRBC suspension was added in each well. The serum was not added in control well the plates were incubated at 370C for about one hrs. The reciprocal of highest dilution showing 50% agglutination was expressed as titer (n) the titer was transformed into log 2 for further analyses.
Material and methods
In the present investigation, chicks belonging to the selected lines of native chickens were used for the study of immunological traits viz., response to sheep RBCs, IgG concentrations. The statistical analysis was performed by using SPSS10 software for evaluating various parameters.
Experimental Animals -Birds and their genetic background
Approximate 100 chickens of native randomly selected from a flok of 300 birds being maintained at experimental farm of CARI, Izatnagar, India as closed flock. Screenings of population for SRBC response in each of the above was used for current study.
Sheep
Healthy Muzaffarnagari breed of sheep maintained at Sheep and Goat farm of Livestock Production Research Centre, Indian Veterinary Research Institute, Izatnagar were used for collection of blood from the Jugular vein to prepare SRBC suspension.
Assaying of Immunological traits
The immunocompetence status of the birds can be assessed by analyzing various components of immune system. A few important facets of immune responses were evaluated in this investigation. Total IgG antibody titre and humoral immune response manifested by its components like antibody titer against an antigen were estimated.
Humoral Response to Sheep RBC
Preparation of Sheep RBC antigen
Approximately 10 ml of heparinized (20 IU/ml) blood was collected from jugular vein of healthy sheep. It was centrifuged at 40C 120C on 3-5 thousand rpm. For 10 minutes to settle down the RBCs. The RBCs were then washed thrice with PBS/NSS by mixing and centrifuging it to remove other serum components. Finally, 1% sheep RBC suspension was prepared by mixing 1 ml of packed sheep RBCs and 99 ml of PBS which was then used for injection in the birds.
Administration of antigen
0.1 ml of 1% sheep RBC suspension was injected into the jugular vein of each bird with tuberculin syringe. Jugular vein was the choice of injection as it led to minimum bleeding in comparison to other veins like brachial vein etc.
Harvesting of immune sera from SRBC sensitized birds
Two ml of blood was collected in sterilized glass tubes at 5 days post immunization (5 dpi) and allowed to clot for 2-3 hours at 370C. The hyper immune sera oozed out of the clot or the clot was broken. Sera were collected by centrifuging the tube at 1000 rpm for 3-4 min. and stored at -200C till further analysis.
Estimation of antibody titer against sheep RBCs
The antibody response to SRBC was assessed using haemagglutination test as mentioned below-
The test was performed in round bottom (U shaped) micro titer plates.50 ml of phosphate buffered saline (PBS) was added in each well. Then, 50 ml of serum was added in first well of each row except the last row where 50 ml of PBS was added, that acted as control. After thorough mixing thoroughly, the sera were two fold serially diluted by taking 50 ml from each of the well and adding it to the subsequent wells, mixing thoroughly and it was continued like this till last column, which was discarded. Equal volume (50 ml) of 1% SRBC suspension was added in all the wells and was thoroughly mixed with sera samples. The plates were then incubated at 37 0C for 1 hour. The highest dilution that gives complete agglutination (button shaped clumping of RBCs indicated haemagglutination reaction) and it was recorded as titer and was expressed as log2n.
Estimation of serum IgG by Single Radial Immuno-diffusion (SRID) Assay
Chicken serum IgG neutralizes the anti chicken IgG Agarose gel was used as a solidifying base to assay IgG concentrations through Single Radial Immunodiffusion (SRID) assay. The procedure is briefed below.
Clean and sterilized glass plate was placed on leveled horizontal surface. The edges prepared with glass strips at the borders were sealed with agar. About 50 ml of 0.1 M Tris – HCl was divided equally into two halves. In first half, of 3% Agarose concentration was added 3% (w/v) and boiled. In second half, 1.750 ml of anti-chicken IgG (Sigma, USA) was added and after thorough mixing, it was kept at 50 0C in a water bath. The temperature of first half (boiled and cooled) was brought down to about 50 0C and second half was mixed. The whole content was poured on to the glass plate. The gel was allowed to solidify for 1-2 h. Then wells were punched at a distance of 1 cm with the help of well-cutter. The standards of IgG (Sigma, USA) were loaded in different concentration in the wells to plot standard curve. 5 ml of unknown sera were diluted to 4 times with 0.1 M Tris and then loaded the wells. The plate was incubated at 37 0C for 24 h in humid chamber. The diameters of the ring around standard as well as unknown samples were measured with the help of Digital Vernier Calipers. The serum IgG concentrations in unknown samples were determined with the help of regression equation obtained by plotting log2 concentrations of IgG standards against diameter of the precipitation ring as mentioned in section 2for serum lysozyme.
Statistical analysis
The data generated on immunological traits were analyzed by LS ANOVA using following Statistical model: –
Y= µ + Li + Li:Sij + Hk + Sxl + eijklm
Where, Y = value of a trait measured on ijklmth individual
µ = Overall mean
Li = Effect of line (i = 1, 2)
Li:Sj = Random effect of jth sire in ith line
Hk = effect of kth hatch
Sxl = Effect of lth sex (l = 1, 2)
eijklm = random error associated with mean ‘0’ and variance σ2
Results and Discussion
The observed results are listed in the following tables and graphs by using SPSS10 software of data Analysis for evaluating various statistical parameters of native Chickens.
The Average means for HA was observed (7.74±0.109) and IgG (1.94±0.72) in native breed. Sex wise HA results showed that male have higher mean (7.925 ± 0.1463) than females (7.57± 0.161) for HA while the same was also estimated for IgG by having mean (2.1194±8.9) and female (1.77±8.18). (Table No.1&2).
Table 1: Mean for HA and IgG Sex wise of Native Chicken.
| SEX | Mean | HA | IGG |
|
Male |
7.9254 | 2.1194 | |
| Std. Error of Mean | .1463 | 8.902 | |
| Std. Deviation | 1.1974 | .7287 | |
|
Female |
Mean | 7.5714 | 1.7714 |
| Std. Error of Mean | .1610 | 8.183 | |
| Std. Deviation | 1.3467 | .6846 | |
|
Total |
Mean | 7.7445 | 1.9416 |
| Std. Error of Mean | .1097 | 6.196 | |
| Std. Deviation | 1.2835 | .7252 |
Table 2: Descriptive One way Anova for HA and IgG Traits.
|
Traits |
Sex |
N |
Mean |
Std. Deviation |
Std. Error |
95% Confidence Interval for Mean
|
|
| HA | Male | 67 | 7.9254 | 1.1974 | .1463 | 7.6333 | 8.2174 |
| Female | 70 | 7.5714 | 1.3467 | .1610 | 7.2503 | 7.8925 | |
| Total | 137 | 7.7445 | 1.2835 | .1097 | 7.5277 | 7.9614 | |
| IGG | Male | 67 | 2.1194 | .7287 | 8.902 | 1.9417 | 2.2971 |
| Female | 70 | 1.7714 | .6846 | 8.183 | 1.6082 | 1.9347 | |
| Total | 137 | 1.9416 | .7252 | 6.196 | 1.8191 | 2.0641 | |
One way Anova recorded significant between the groups of male and females having less than 0.05 values which is recommended for significant level. The square mean value was observed for HA (4.3) & IgG (4.145) between the groups, while with in the group was observed as (1.628) for HA and IgG (0.499). (Table No.4)
Test of Homogeneity of Variances was recorded as (0.39) for HA which was significant and IgG level (0.937). (Table No.3).
Table 3: Test of Homogeneity of Variances for HA and IgG Traits
| Traits | Levene Statistic | df1 | df2 | Sig. |
| HA | 4.363 | 1 | 135 | .039 |
| IGG | .006 | 1 | 135 | .937 |
Table 4: One Way ANOVA Analysis for HA and IgG Traits.
| Traits | Type | Sum of Squares | df | Mean Square | F | Sig. |
| HA | Between Groups | 4.289 | 1 | 4.289 | 2.634 | .107 |
| Within Groups | 219.770 | 135 | 1.628 | |||
| Total | 224.058 | 136 | ||||
| IGG | Between Groups | 4.145 | 1 | 4.145 | 8.304 | .005 |
| Within Groups | 67.388 | 135 | .499 | |||
| Total | 71.533 | 136 |
A Graph was plotted between Mean and Sex for determining the level of immunity as follows:
The native breeds have unique alleles in their genetic constitution being reared in agro-climatic conditions for their well adaptability in open environment. Result showed that native breed was estimated for HA was observed (7.74±0.109) and IgG (1.94±0.72) which was higher being a native breed and it was preserved in open environment under extreme climatic conditions by which their immunity levels has increased. A mean titer of 6.289±0.246 in SDL broiler chickens. Aseel breed of native chicken demonstrated higher HA titer than most of the other breeds20. The wide variations observed in the present study might be due to the reason that no artificial selection has been applied on this breed of chicken for immune response or production traits.
Sex wise HA results showed that male have higher mean (7.925 ± 0.1463) than females (7.57± 0.161) for HA while the same was also estimated for IgG by having mean (2.1194±8.9) and female (1.77±8.18). This difference might be due to male and female antibody titers being genetically different traits, due to the genes being located on the sex chromosome. This possibility can be studied by estimating the genetic correlation between male and female antibody titers. The overall male has better immunity level then females. The least squares mean of HA titer as 12.38±0.60 in Aseel native chicken. In males, the average was 12.80±0.74 whereas in case of females, it was 12.35±0.62.
Besides, genetic factors other factors have also been reported to influence the response to sheep RBCs 20, 21. One way Anova recorded significant between the groups of male and females having less than 0.05 values which are recommended for significant level. The square mean value was observed for IgG (4.145) between the groups, while with in the group was observed as (0.499). Serum IgG is the most abundant antibody and constitutes approximately 80% of the total immunoglobulin. The bird’s ability to mount antibody responses to other antigens is primarily revealed by serum IgG concentration. The average serum IgG concentration was 6.987±6.57 it was higher in males (7.409±8.13) than females (6.533±10.20). The serum IgG concentrations in broiler & indigenous birds were reported to be 8.01±0.4 and 10.01±0.4 mg/ml, respectively21.
Factors Affecting Immunity
There are several factors which may effect immunity level of an host are as follows –
Age of the birds had no significant (P>0.05) effect on serum IgG level. However, older birds revealed somewhat higher values. Sex of the birds had no significant (P>0.05) effect on serum IgG level. However, male birds revealed higher values. Singh (2005) reported non-significant effect of sex on IgG concentrations. The least squares means for males and females were 59.09±7.69 and 52.08±7.74 mg/ml in WLH, respectively.
The influence of age was significant (P<0.05) on HA titer (Table 4.1.1), older birds had higher mean antibody titer (13.0±2.13) than younger birds (10.88±0.54). This results has also reported effect of age on HA titer19. The influence of sex on HA titre was statistically non-significant although males had higher mean antibody titer (12.80±0.74) than females (11.96±0.64). The finding is comparable to those reported by in different genetic stock20, 21, 22.
Sex had non-significant effect on HA titre in guinea fowl too21. However significant sex differences (females having more titer than males) have been reported in 9th generation of divergent selection based on SRBC response. Sexual dimorphism was attributed to the effect of sex hormone on the thymus and the immune cells as seen in mammals17. Some outcome we get, Native Breed of chicken demonstrated relatively higher humoral response to Sheep erythrocytes than exotic chicken breed, Serum IgG level was also relatively significant in native chicken, Male showed higher immune response then the females.
 |
Figure 1: Mean for HA sex wise. Click here to View figure |
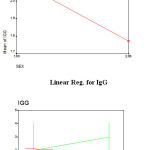 |
Figure 2: Mean for IgG
|
Acknowledgments
I am highly thanks full to my technical staff.
References
- Cheng, H.W., Eicher, S.D., Chen,Y., Singleton P., Muirt, W.M. Effect of genetic selection for group productivity and longevity on immunological and haematological parameters of chickens. Sci. 2001 ; 80 : 1079–1086.
- Ewald, S.J., Lien Y.Y., L. Li, Johnson, L.W. B-haplotype control of CD4/CD8 subsets and TCR V beta usage in chicken T lymphocytes. Immunol. Immunopathol. 1996 ; 53 : 285–301.
- Gavora, J.S. Genetic control of disease resistance in poultry. In:Manipulation of the Avian Genome (Ed. by R.J. Etches and A.M.Varrinder, Gibbins), CRC Press, Boca Raton, FL. 1993 ; 231- 41.
- Collisson, E.W., Pei, J., Dzielawa J., Seo, S.H. Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev.Comp. Immunol. 2000 ; 24 : 187–200.
- Hunt, H.D., Lupiani, B., Miller, M.M., Gimeno, I., Lee, L.F., Parcells, M.S. Marek‘s disease virus down-regulates surface expression of MHC (B Complex) Class I (BF) glycoproteins during active but not latent infection of chicken cells. Viro. 2001 ; 282 : 198–205.
- Juul-Madsen, H.R., Nielsen, O.L., Krogh-Maibom, T., Rontved, C.M., Dalgaard, T.S., Bumstead, N., Jorgensen, P.H. Major Histocompatibility complex-linked immune response of young chickens vaccinated with an attenuated live infectious bursal disease virus vaccine followed by an infection. Poult. Sci. 2002 ; 81 : 649–656.
- Loa, C.C., Lin, T.L., Wu, C.C., Bryan, T., Thacker, H.L., Hooper, T., Schrader, D. Humoral and cellular immune responses in turkey poultsinfected with turkey corona virus. Poult. Sci. 2001 ; 80 : 1416–1424.
- Mazanec, M.B., Kaetzel, C.S., Lamm, M.E., Fletcher, D., Nedrud, J.G. Intracellular neutralization of virus by immunoglobulin antibodies. Proc. Natl. Acad. Sci. 1992 ; 89 : 6901-6905.
- Knowles, S.C., Nakagawa, L.S., Sheldon, B.C. Elevated reproductive effort increases blood parasitaemia and decreases immune function in birds: a metaregression approach. Func. Ecolo. 2009 ; 23 : 405- 415.
- Verhulst, S., Riedstra, B., Wiersma, P. Brood size and immunity costs in zebra finches Taeniopygia guttata. J. of Avi. Biol. 2005 ; 36 : 22-30.
- Cheng, S., Lamonth, S.J. Genetic analysis of immunocompetence measure in a White Leghorn Chicken line. Poultry Sci. 1988a ; 67 (7) : 989-995.
- Sarker, N., Tsudzuki, M., Nishibori, M., Yasue, H., Yamamoto, Y. Cell-mediated and humoral immunity and phagocyticability in chicken lines divergently selected for serum immunoglobulin M and G levels. Pou. Sci. 2000 ; 79 : 1705.
- Li, Z., Nestor, K.E., Saif, Y.M., Anderson, J.W., Patterson, R.A. Effect of selection for increased body weight in turkey on lymphoid organ weights, phagocytises, and antibody responses to fowl cholera and Newcastle disease-inactivated vaccines. Poult. Sci. 2001 ; 80 : 689-694.
- Yunis, R., Ben-David, A., Heller, E.D., Cahaner, A. Antibody responses and morbidity following infection with infectious bronchitis virus ad challenge with Escherichia coli in lines divergently selected on antibody response. Pou. Sci. 2002 ; 81 : 149-159.
- Pinard, M.H., van Arendonk, J.A.M., Nieuwland, M.G.B., van der Zijpp, A.J. Divergent selection for immune responsiveness in chickens: Estimation of Realized heritability with an animal model. J. Anim. Sci. 1992 ; 70 : 2986–2993.
- Gross, W.B., Siegel, P.B., Pierson, F.W. Effects of genetic selection for high or low antibody response on resistance to a variety of disease challenges and the relationship of resource allocation. Avian Dis. 2002 ; 46 : 1007–1010.
- Boa-Amponsem, K., Price, S.E.H., Dunnington, E.A., Siegel, P.B. Effect of route of inoculation on humoral immune response of White Leghorn chickens selected for high or low antibody response to sheep red blood cells. Poult. Sci. 2001 ; 80 : 1073–1078.
- Kundu, A., Singh, D.P., Mohapatra, S.C., Dash, B.B., Moudgal, R.P., Bisht, G.S. Antibody response to sheep erythrocytes in Indian native vis-à-vis imported breeds of chickens. Br. Poultry Sci. 1999 ; 40 : 40-43
- Ubosi, C.O., Dunnington, E.A., Gross, W.B., Siegel, P.B. Divergent selection of chickens for antibody response to sheep erythrocytes kinetics of primary and secondary immunizations. Avian Dis. 1985 ; 29 : 347–355.
- Sivaraman, G.K., Kumar, S., Saxena, V.K., Singh, N.S., Shiva kumar, B.M., Muthukumar, S.P. Genetics of Immunocompetenttraits in a Synthetic Broiler Dam Line. Bri. Pou. Sci. 2005 ; 46 : 169-74.
- Saxena, V.K., Singh, H., Pa, S.K., Kumar, S. Genetic studies on primary response to sheep erythrocytes in guinea fowl. Bri. Pou. Sci. 1997 ; 38 : 156-58.
- Singh, A.P. Interleukin receptor genes polymorphism in specialized layer chicken lines. M.V.Sc. Thesis, Indian Veterinary Research Institute, Izatnagar, Deemed University, India. 2005

