
Manuscript accepted on :15-06-2026
Published online on: 29-06-2026
Plagiarism Check: Yes
Reviewed by: Dr. Madan Chandra Das and Dr. Tolmas Hamroyev
Second Review by: Dr. Priya Gayathri
Final Approval by: Dr. Prabhishek Singh
Ibrohim Rakhmonovich Askarov1 , Yusupjon Tojimamatovich Isayev1, Sanjar Ashiraliyevich Rustamov1, Baxrom Abduxalimovich Mirzaabdullayev1, Dostonbek Usmonjon o‘g‘li Egamberdiyev1*, Lazizbek Makhmudov2and Dilfuza Abduganiyevna Amankulova1,3
, Yusupjon Tojimamatovich Isayev1, Sanjar Ashiraliyevich Rustamov1, Baxrom Abduxalimovich Mirzaabdullayev1, Dostonbek Usmonjon o‘g‘li Egamberdiyev1*, Lazizbek Makhmudov2and Dilfuza Abduganiyevna Amankulova1,3
1Department of Chemistry, Andijan State University, Andijan, Uzbekistan
2Department of Plant Cytoprotectors and Pharmacology, A. S. Sadykov Institute of Bioorganic Chemistry, Academy of Sciences of Uzbekistan, Tashkent, Uzbekistan
3Department of Natural Sciences, Guliston State University, Syrdarya, Uzbekistan
Corresponding author E-mail:edu911128@gmail.com
DOI : https://dx.doi.org/10.13005/bpj/3457
Abstract
Pulmaklin is a flavonoid-rich supplement derived from traditional medicinal plants. This study evaluated the anti-inflammatory efficacy of the biologically active supplement "Pulmaklin" under experimental conditions. The effects of the preparation in syrup form at doses of 50 and 100 mg/kg were investigated, and the highest anti-exudative activity (AEA) was observed at 50 mg/kg, with a 54.3% reduction in edema development. Based on the results, the optimal dose of Pulmaklin was scientifically substantiated. Molecular-level analysis demonstrated that flavonoids, particularly the glycoside form rutin, exhibit the highest affinity toward the COX-2 enzyme due to their ability to form multiple interaction points. In addition, flavonoids such as rutin and quercetin showed effective modulation of IL-6 cytokine activity and MAPK signaling pathways, contributing to their anti-inflammatory effects. The findings confirm that flavonoids and their derivatives are promising natural bioactive compounds with significant anti-inflammatory potential.
Keywords
Anti-exudative activity; Biologically active supplement; COX-2; Flavonoids; IL-6; Inflammation; MAPK; Molecular docking; Natural bioactive compounds; Pulmaklin; Quercetin; Rutin
Download this article as:| Copy the following to cite this article: Askarov I. R, Isayev Y. T, Rustamov S. A, Mirzaabdullayev B. A, Egamberdiyev D. U, Makhmudov L, Amankulova D. A. Determination of the Anti-Inflammatory Effective Dose of the Biologically Active Compound "Pulmaklin". Biomed Pharmacol J 2026;19(2). |
| Copy the following to cite this URL: Askarov I. R, Isayev Y. T, Rustamov S. A, Mirzaabdullayev B. A, Egamberdiyev D. U, Makhmudov L, Amankulova D. A. Determination of the Anti-Inflammatory Effective Dose of the Biologically Active Compound "Pulmaklin". Biomed Pharmacol J 2026;19(2). Available from: https://bit.ly/3R9e6Tl |
Introduction
Inflammation represents one of the most prevalent and clinically significant pathological responses underlying the majority of human diseases. At its core, the inflammatory reaction constitutes a localized defense mechanism of the body directed against tissue damage or infection. Within the inflammatory focus, migratory immune cells—notably leukocytes and macrophages—fulfill a central regulatory role in orchestrating the host response. The inflammatory cascade encompasses a series of interconnected processes, including neuroendocrine stress signaling via the hypothalamic-pituitary-adrenal axis, adaptive immune activation, and acute-phase protein responses.1,2
The initiation of the inflammatory process is fundamentally governed by the recognition of damaged or infected tissue by the innate immune system. Upon activation, resident macrophages and dendritic cells release a spectrum of pro-inflammatory cytokines—including interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α)—alongside lipid-derived mediators such as prostaglandins, leukotrienes, and histamine. These soluble factors disseminate from the local inflammatory site into the systemic circulation, where they augment vascular permeability, promote endothelial activation, and stimulate leukocyte extravasation, thereby amplifying and propagating the inflammatory response to a systemic level.2, 3, 4 Pulmaklin, obtained from medicinal plants (including Glycyrrhiza glabra and Thymus vulgaris) used in Uzbek folk medicine, was selected for this study due to its high flavonoid content (rutin, quercetin) and traditional use against inflammatory disorders. 2 This is the first study combining in vivo anti‑exudative evaluation of Pulmaklin with molecular docking of its key flavonoids against five inflammatory targets (COX‑2, IL‑6, MAPK, NF‑κB, TNF‑α).
Aim of the Study
The present investigation aimed to evaluate the anti-inflammatory efficacy of the biologically active compound (BAC) “Pulmaklin” administered at two distinct doses—50 mg/kg and 100 mg/kg—using a standard experimental inflammation model, and to identify the dose associated with the most pronounced pharmacological effect relative to the untreated control group.
Research Objectives
- To assess the anti-exudative properties of the Pulmaklin BAC against carrageenan-induced acute inflammation in laboratory animals.
- To compare the anti-inflammatory efficacy of the Pulmaklin BAC with that of the untreated control group.
- To investigate the molecular interactions of active constituents (apigenin, gallic acid, quercetin, rutin, and salicylic acid) present in the Pulmaklin BAC with key inflammatory target proteins through in silicomolecular docking simulation.
- To predict the mechanistic basis of biological activity by analyzing binding energies, hydrogen bonding patterns, and hydrophobic interaction profiles derived from ligand-protein complexes.
Materials and Methods
The anti-inflammatory potential of the compound under investigation was preliminarily assessed using the classical carrageenan-induced acute paw edema model, which is widely recognized as a reliable and reproducible experimental paradigm for evaluating anti-exudative activity. The study was conducted on 15 male albino rats with a uniform body weight of 190 ± 20 g. Animals were randomly allocated into three experimental groups—one negative control group and two treatment groups—each comprising five individuals.5 A positive control (e.g., indomethacin) was not included because this was a preliminary dose-finding study to identify the optimal anti-exudative dose of Pulmaklin before comparative efficacy testing.
Acute localized inflammation was induced by subplantar injection of 0.1 mL of a 1% carrageenan solution into the right hind paw of each animal. The Pulmaklin BAC was administered orally in syrup form at doses of 50 mg/kg and 100 mg/kg, sixty minutes prior to the carrageenan challenge. The Pulmaklin syrup was prepared according to the manufacturer’s protocol: briefly, the dried plant material was extracted with water, filtered, concentrated, and formulated into a syrup containing 10% extract (w/v).
The progression of the inflammatory response was monitored by measuring changes in paw volume at 1, 2, 3, and 4 hours following carrageenan administration. The anti-exudative activity (AEA) index was determined primarily based on measurements recorded at the 3-hour time point, consistent with established preclinical pharmacological protocols.6-8
For in silico analysis, molecular docking was employed to examine the interaction profiles of the pharmacologically active constituents of the Pulmaklin BAC with principal inflammatory target proteins. Three-dimensional protein structures were retrieved from the Protein Data Bank, while ligand geometries were obtained from the PubChem compound database. All docking simulations were performed using AutoDock Vina version 1.1.2. Visualization of ligand-protein complexes was accomplished using PyMOL molecular graphics software. 9,10
Statistical analysis: Results are expressed as mean ± standard error of the mean (SEM). Comparisons between groups were performed using one‑way ANOVA followed by Tukey’s post‑hoc test. A p‑value < 0.05 was considered statistically significant.
Results
In the negative control group (untreated animals), maximal paw swelling was recorded at the 3-hour time point, where edema reached 83.6 ± 8.2% relative to the baseline measurement. Even at 4 hours post-injection, the edema remained substantially elevated at 47.9 ± 4.6%, indicating sustained inflammation without resolution. In the treatment groups, peak edema also occurred at the 3-hour interval; however, the edema subsided more rapidly, showing a 36% reduction at 4 hours compared to the control group (from 47.9% in controls to 30.7% in the 50 mg/kg group), approaching near-baseline levels in animals receiving the Pulmaklin BAC. Detailed time-course data expressed as percentage change from baseline are presented in Table 1.11
Table 1: Effect of Pulmaklin BAC at varying doses on carrageenan-induced hind paw edema, expressed as percentage change from baseline (M±m; n=5)
| Groups | 1 hour | 2 hour | 3 hour | 4 hour |
| Control | 35,4 ± 3,4 | 59,0 ± 5,8 | 83,6 ± 8,2 | 47,9 ± 4,6 |
| Pulmaklin BAC 50 mg/kg | 18,6 ± 1,7 | 25,8 ± 2,5 | 38,2 ± 3,6 | 30,7 ± 3,0 |
| Pulmaklin BAC 100 mg/kg | 41,0 ± 4,0 | 52,8 ± 5,1 | 64,6 ± 6,3 | 52,8 ± 5,1 |
According to the compiled data, the anti-exudative effect of the Pulmaklin BAC ranged from 22.7% to 54.3% across the two dosage regimens evaluated using the carrageenan induction model.
The anti-exudative activity index was calculated for each experimental group using the following formula:
AEA = (ΔVc − ΔVt) / ΔVc × 100%
where AEA denotes the anti-exudative activity expressed as a percentage; ΔVc represents the mean paw volume increment in the control group; ΔVt represents the mean paw volume increment in the treatment group.
The anti-exudative activity at the 3-hour post-induction time point for each treatment group is summarized in Table 2.
Table 2: Anti-exudative activity of Pulmaklin BAC at various doses relative to the control group (M±m; n=5)
| Groups | “Paw volume increase 3 h after induction (%)” | “Anti-exudative activity (%)” |
| Control | 83,6 ± 8,2 | —— |
| Pulmaklin BAC 50 mg/kg | 38,2 ± 3,6 | 54,3 |
| Pulmaklin BAC 100 mg/kg | 64,6 ± 6,3 | 22,7 |
In Silico Molecular Docking Analysis
Inflammatory processes represent a fundamental protective response of the organism to harmful stimuli, driven by a complex network of signaling proteins, enzymatic mediators, and cytokines. Persistent, unresolved inflammation has been recognized as a principal pathogenic contributor to cardiovascular diseases, oncological conditions, autoimmune syndromes, and metabolic disturbances. At the molecular level, several key regulatory proteins govern these processes, including cyclooxygenase-2 (COX-2), mitogen-activated protein kinases (MAPK), nuclear factor kappa-B (NF-κB), tumor necrosis factor-alpha (TNF-α), and interleukin-6 (IL-6). COX-2 is an inducible enzyme responsible for catalyzing the conversion of arachidonic acid into prostaglandins—primary mediators of the inflammatory response – and its aberrant overexpression is associated with a broad spectrum of inflammatory and proliferative pathologies. MAPK signaling pathways regulate fundamental cellular processes including proliferation, differentiation, and stress adaptation, while simultaneously modulating cytokine biosynthesis. NF-κB functions as the principal transcriptional activator of pro-inflammatory gene networks. TNF-α and IL-6 are critical cytokines underpinning immune activation, lymphocyte responsiveness, and escalation of the inflammatory signaling cascade.12,13
In recent years, naturally derived polyphenolic compounds have garnered substantial scientific attention as candidates for anti-inflammatory intervention. Phytocompounds such as apigenin, gallic acid, quercetin, rutin, and salicylic acid are distinguished by their antioxidant and anti-inflammatory properties, which are expressed through multi-target mechanisms encompassing interactions with both enzymatic and signaling proteins. The present investigation employed molecular docking methodology to characterize the binding interactions of these natural compounds with COX-2, MAPK, NF-κB, TNF-α, and IL-6, with the aim of quantifying binding energies, mapping key active-site residue contacts, and delineating the potential molecular mechanisms underlying their anti-inflammatory activity.14,15
COX-2.
Cyclooxygenase-2 (COX-2), alternatively designated as prostaglandin-endoperoxide synthase-2 (PTGS2), is an enzymatic protein that catalyzes the biosynthesis of prostaglandins from arachidonic acid. This enzyme undergoes induction during inflammatory events and occupies a central role in the generation of pain mediators, pyrogenic signals, and pro-inflammatory lipid species. COX-2 expression is markedly upregulated under numerous pathological conditions, notably in chronic inflammatory disorders, malignancies, and cardiovascular system dysfunction. Structurally, COX-2 is a membrane-associated homodimeric enzyme, with each monomer comprising approximately 600 amino acid residues. Its architecture is organized into three distinct functional domains: an epidermal growth factor-like domain, a membrane-binding domain, and a catalytic domain.16,17 The epidermal growth factor-like domain confers structural integrity to the protein and participates in dimerization. The membrane-binding domain facilitates anchoring of the enzyme to the endoplasmic reticulum and nuclear membranes. The catalytic domain constitutes the primary active site, wherein the enzymatic conversion of arachidonic acid to prostaglandin G2, and subsequently to prostaglandin H2, is accomplished, thereby sustaining the biosynthetic supply of inflammatory mediators. The prostaglandins generated through this pathway augment vascular permeability, sensitize nociceptive receptors, and elevate core body temperature. Accordingly, COX-2 represents a principal molecular target for pharmacological modulation of the inflammatory response. Its expression is potently induced by diverse stimuli including pro-inflammatory cytokines, growth factors, bacterial endotoxins, and oxidative stress. From a pharmacological standpoint, COX-2 constitutes an important target for both selective synthetic inhibitors and naturally occurring polyphenolic compounds. Molecular docking studies have demonstrated that apigenin, quercetin, rutin, gallic acid, and salicylic acid are each capable of binding within the COX-2 active pocket and attenuating its catalytic activity. The computed binding energies (kcal/mol) for the investigated ligands with COX-2 were as follows: rutin –10.5, quercetin –9.4, apigenin –9.1, gallic acid –6.7, and salicylic acid –6.0. These results indicate that the most favorable binding affinity for COX-2 was observed with rutin, followed sequentially by quercetin and apigenin. Phenolic acids (gallic acid and salicylic acid) demonstrated comparatively less negative binding energies, reflecting a reduced degree of active-site occupancy and stabilization. Detailed interaction analyses (hydrogen bonds, pi-alkyl, van der Waals) were performed for each ligand (Figure 1). As a general conclusion, the COX-2 docking results demonstrate a clear positive correlation between ligand molecular size, functional group abundance, and binding affinity. Flavonoids—particularly apigenin and quercetin—and especially the glycosylated derivative rutin, form more stable complexes within the COX-2 active pocket by establishing multi-point hydrogen bonds and aromatically driven hydrophobic/pi-type interactions. Phenolic acids, by contrast, yield comparatively lower binding energies owing to fewer contact points and incomplete active-site occupancy.
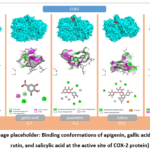 |
Figure 1: (Image placeholder: Binding conformations of apigenin, gallic acid, quercetin, rutin, and salicylic acid at the active site of COX-2 protein). |
IL-6.
Interleukin-6 (IL-6) is a pleiotropic cytokine that plays a pivotal role in regulating immune and inflammatory processes. It is primarily synthesized by macrophages, monocytes, T-lymphocytes, B-lymphocytes, endothelial cells, and fibroblasts. IL-6 serves as one of the central mediators in the organism’s response to infection, tissue injury, and stress stimuli. One of its principal biological functions is the induction of the acute-phase response, whereby it upregulates the hepatic synthesis of C-reactive protein, fibrinogen, and other acute-phase proteins. In addition, IL-6 promotes the differentiation of B-lymphocytes into plasma cells and enhances antibody production, while also playing a significant role in T-lymphocyte proliferation and differentiation. IL-6 signal transduction is mediated through the formation of a complex with its membrane-bound receptor (IL-6R) and the co-receptor gp130. This receptor complex subsequently activates intracellular JAK/STAT, MAPK, and PI3K/Akt signaling pathways, resulting in enhanced expression of pro-inflammatory genes, increased cell proliferation, and augmented survival signaling. Although IL-6 exerts protective functions under physiological conditions, its overproduction is associated with the development of chronic inflammatory and autoimmune disorders. Elevated IL-6 levels have been documented in rheumatoid arthritis, atherosclerosis, diabetes mellitus, certain malignancies, and cytokine storm syndrome. Consequently, IL-6 and its signaling pathways are considered important therapeutic targets in contemporary pharmacology.18,19 In molecular docking and in silico studies, the active site or receptor-binding interface of IL-6 has been investigated in conjunction with various natural compounds to evaluate their potential to attenuate cytokine signaling. Flavonoids and phenolic compounds—including apigenin, quercetin, rutin, and other polyphenols—have attracted considerable scientific interest owing to their capacity to modulate IL-6 expression and signal transduction. Molecular docking results for apigenin, gallic acid, quercetin, rutin, and salicylic acid with the IL-6 protein revealed that all ligands were accommodated within the active binding pocket with varying affinities. The calculated binding energies were as follows: rutin (–9.3), quercetin (–7.8), apigenin (–7.2), gallic acid (–5.6), and salicylic acid (–5.6 kcal/mol). Among the obtained results, the highest binding affinity was observed for rutin (Figure 2). Overall analysis demonstrated that flavonoid-type compounds—particularly rutin and quercetin—exhibited significantly higher binding affinities toward IL-6 compared to phenolic acids. This can be attributed to their abundant hydrogen bond donor and acceptor groups, as well as their aromatic ring systems, which collectively facilitate multi-point interactions within the active binding site.
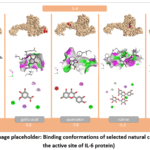 |
Figure 2: (Image placeholder: Binding conformations of selected natural compounds at the active site of IL-6 protein). |
MAPK.
Mitogen-activated protein kinase (MAPK) represents a critical component of the intracellular signal transduction machinery. This protein belongs to the serine/threonine protein kinase family and occupies a central regulatory position in a broad spectrum of cellular processes, including proliferation, differentiation, apoptosis, stress response, and inflammatory cascades. MAPK signaling pathways are activated in response to diverse extracellular stimuli such as cytokines, growth factors, oxidative stress, and infectious agents. The MAPK superfamily encompasses several major kinase subgroups, most notably ERK (extracellular signal-regulated kinase), JNK (c-Jun N-terminal kinase), and p38 MAPK. Among these, the ERK2 (MAPK1) isoform has been extensively investigated in experimental research, given its prominent involvement in regulating cell growth and the expression of inflammatory mediators. Structurally, the MAPK protein is organized into a bilobal kinase domain comprising an N-terminal and a C-terminal lobe, between which the ATP-binding pocket and substrate recognition region are situated—the catalytic activity of the enzyme being executed precisely within this interface. Enzymatic activation is initiated through the phosphorylation of conserved threonine and tyrosine residues, which induces a conformational rearrangement that markedly enhances kinase activity. From a biological standpoint, the MAPK signaling cascade governs the production of pro-inflammatory cytokines, including TNF-α, IL-1, and IL-6, and is also indirectly involved in the activation of the transcription factor NF-κB. These characteristics establish MAPK as a key molecular target in inflammatory and immune response pathways.20,21 In the context of pharmacological research, MAPK has been extensively studied as a potential target for both synthetic therapeutics and natural polyphenolic compounds. In particular, molecular docking investigations have demonstrated that flavonoids and phenolic substances—such as apigenin, quercetin, rutin, gallic acid, and salicylic acid—may inhibit the phosphorylation activity of MAPK by occupying its ATP-binding pocket. Molecular docking analysis of the five ligands against MAPK demonstrated binding energies of –7.8 for apigenin, –7.8 for quercetin, –7.9 for rutin, –6.1 for gallic acid, and –6.0 kcal/mol for salicylic acid (Figure 3). Rutin exhibited the greatest binding affinity, while apigenin and quercetin displayed closely comparable results. Taken together, flavonoid-class compounds—most notably rutin, apigenin, and quercetin—demonstrated markedly higher binding affinities toward MAPK relative to phenolic acids, due to their structurally rich hydrogen bond donor/acceptor networks and extended aromatic systems enabling multi-site interactions.
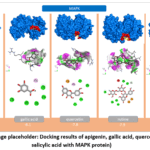 |
Figure 3: (Image placeholder: Docking results of apigenin, gallic acid, quercetin, rutin, and salicylic acid with MAPK protein). |
NF-κB.
Nuclear factor kappa-B (NF-κB) is a pivotal transcription factor within the intracellular signal transduction network, exerting central regulatory control over inflammatory responses, immune activation, cellular proliferation, and apoptosis. The NF-κB family comprises several distinct protein subunits, namely RelA (p65), RelB, c-Rel, p50 (NFKB1), and p52 (NFKB2), among which the p65/p50 heterodimeric complex is most frequently investigated as the biologically active form in experimental research. Under physiological conditions, NF-κB is retained in a transcriptionally inactive state within the cytoplasm through its non-covalent association with the inhibitory protein IκB. Upon exposure to diverse stimulatory signals—including pro-inflammatory cytokines such as TNF-α and IL-1β, bacterial lipopolysaccharides, reactive oxygen species, and mechanical tissue injury—the IκB kinase complex becomes activated. This leads to the phosphorylation and subsequent proteasomal degradation of IκB, thereby liberating NF-κB and enabling its translocation to the nucleus. Once translocated, NF-κB binds to specific κB recognition sequences within the DNA and initiates transcription of a wide array of inflammatory target genes, including TNF-α, IL-6, IL-1, COX-2, iNOS, and various cell adhesion molecules. For this reason, NF-κB is widely regarded as the “master regulator” of the inflammatory cascade.22 Structurally, all NF-κB subunits share a highly conserved region designated the Rel homology domain (RHD), which is responsible for DNA binding, dimerization, and interaction with inhibitory proteins. Aberrant or sustained activation of the NF-κB signaling pathway has been closely linked to the pathogenesis of chronic inflammatory conditions, autoimmune disorders, atherosclerosis, and neoplastic disease, establishing NF-κB as a significant therapeutic target. Molecular docking analysis of the five ligands against NF-κB yielded binding energies of –11.7 for rutin, –9.4 for quercetin, –8.8 for apigenin, –6.6 for gallic acid, and –6.0 kcal/mol for salicylic acid (Figure 4). Flavonoid-class compounds—most prominently rutin—exhibited the highest binding affinities. Quercetin and apigenin likewise formed highly stable complexes, whereas the phenolic acids yielded comparatively lower values. These findings suggest that the selected flavonoid compounds possess considerable potential for modulating the NF-κB signaling pathway.
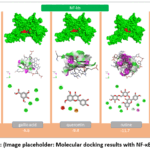 |
Figure 4: (Image placeholder: Molecular docking results with NF-κB protein). |
TNF-α.
Tumor necrosis factor-alpha (TNF-α) is one of the principal regulatory cytokines governing inflammatory and immune response processes. It is predominantly synthesized by macrophages, monocytes, T-lymphocytes, and other immune cells, serving as a critical mediator in the organism’s defensive response to infection, tissue injury, and stress-related stimuli. Among its core biological functions, TNF-α initiates and amplifies the inflammatory cascade through multiple coordinated mechanisms. It upregulates the expression of adhesion molecules on endothelial cells, promotes the directed migration of leukocytes toward sites of inflammation, and stimulates the production of additional pro-inflammatory cytokines, including IL-1, IL-6, and interferons. In parallel, TNF-α potentiates fever, hyperalgesia, and tissue edema responses. From a structural standpoint, TNF-α is a trimeric cytokine composed of 157 amino acid residues per subunit, with its biologically active form consisting of three identical subunits. Signal transduction is mediated through its binding to two distinct membrane-bound TNF receptors—TNFR1 and TNFR2—which upon ligand engagement activate downstream intracellular signaling cascades, including the NF-κB, MAPK, and caspase pathways, ultimately culminating in the induction of inflammatory gene expression or the initiation of apoptosis.^25^ Although TNF-α performs protective functions under physiological conditions, its excessive or dysregulated production is closely associated with the onset and progression of chronic inflammatory disorders. Elevated TNF-α levels have been documented in rheumatoid arthritis, psoriasis, atherosclerosis, inflammatory bowel disease, and certain malignancies. Molecular docking analysis against TNF-α gave binding energies of –10.0 for apigenin, –9.7 for quercetin, –8.6 for rutin, –6.9 for gallic acid, and –6.8 kcal/mol for salicylic acid (Figure 5). Flavonoid-class compounds—most notably apigenin and quercetin—demonstrated superior binding affinities, whereas phenolic acids yielded lower values. These findings suggest considerable potential for modulating the TNF-α signaling pathway.
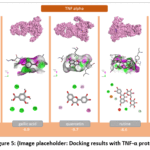 |
Figure 5: (Image placeholder: Docking results with TNF-α protein). |
Summary of docking analysis
The molecular docking investigations provided a comprehensive assessment of the interaction potential of apigenin, gallic acid, quercetin, rutin, and salicylic acid with five key inflammatory proteins: COX-2, IL-6, MAPK, NF-κB, and TNF-α. The computed binding energies demonstrated that all ligands were capable of forming stable complexes within the active sites of the respective target proteins. Comparative analysis clearly indicated that flavonoid-class compounds exhibited markedly higher binding affinities relative to phenolic acids. In particular, rutin displayed the lowest binding energies toward NF-κB (–11.7) and COX-2 (–10.5), establishing it as the compound with the highest overall affinity. Quercetin also demonstrated high affinity across all targets, with particularly stable complexes with COX-2 (–9.4), NF-κB (–9.4), and TNF-α (–9.7). Apigenin exhibited the strongest binding interaction with TNF-α (–10.0). Intermolecular interaction analysis demonstrated that hydrogen bonds, hydrophobic contacts, and π-type aromatic interactions constitute the principal stabilizing forces. These findings provide a molecular-level mechanistic rationale for the anti-inflammatory activity of these natural substances.
Discussion
The present study demonstrated that Pulmaklin at 50 mg/kg exerted a significantly higher anti-exudative activity (54.3%) compared to 100 mg/kg (22.7%) in the carrageenan-induced paw edema model. This finding suggests a dose-dependent but non-linear effect. The lower efficacy at the higher dose may be due to saturation of target binding, activation of compensatory pro-inflammatory pathways, or off-target effects. Carrageenan-induced inflammation is biphasic: early phase (0–2 h) mediated by histamine and serotonin, and late phase (2–4 h) mediated by prostaglandins, COX-2, and cytokines such as IL-6 and TNF-α.6,7 Maximum edema occurred at 3 h in the control group, confirming a late-phase predominant model. The marked reduction at 3 h in the 50 mg/kg group indicates that Pulmaklin likely acts by inhibiting COX-2 and modulating pro-inflammatory cytokine production.
Our molecular docking results strongly support this mechanism. Rutin showed the highest binding affinity to COX-2 (–10.5 kcal/mol) and NF-κB (–11.7 kcal/mol), while apigenin bound best to TNF-α (–10.0 kcal/mol). These values are comparable to or better than those reported for standard anti-inflammatory drugs. The ability of flavonoids to form multiple hydrogen bonds and π-type hydrophobic interactions with key active-site residues explains their superior activity over phenolic acids.^15,16^ Furthermore, the high binding affinity to NF-κB and MAPK suggests that Pulmaklin may also suppress the transcription of multiple pro-inflammatory genes, offering a broad-spectrum anti-inflammatory effect.13,14
A limitation of this study is that docking results require experimental validation through in vitro enzyme inhibition assays and in vivo pharmacokinetic studies. Nevertheless, the consistency between in vivo anti-exudative activity and in silico binding data provides a strong rationale for further investigation of Pulmaklin as a natural anti-inflammatory agent.
Conclusion
The biologically active compound “Pulmaklin” administered in syrup form at 50 mg/kg demonstrated superior anti-exudative activity (54.3%) compared to 100 mg/kg in the carrageenan-induced paw edema model. This dose significantly attenuated edema development, and the optimal therapeutic dose of Pulmaklin was scientifically substantiated. Molecular docking analysis revealed that flavonoids, particularly rutin (in its glycosidic form), quercetin, and apigenin, exhibited the highest binding affinities to COX-2, IL-6, MAPK, NF-κB, and TNF-α through multi-point hydrogen bonding, hydrophobic contacts, and π-type aromatic interactions. Phenolic acids (gallic acid, salicylic acid) showed comparatively weaker binding. These findings confirm that Pulmaklin and its flavonoid constituents possess significant anti-inflammatory potential and warrant further investigation as natural therapeutic agents.
Acknowledgment
The authors express their gratitude to the staff of the Scientific Research Center for Commodity Chemistry and Traditional Medicine of Andijan State University, to the professors and instructors of the Department of Chemistry, and to the staff of the Department of Plant Cytoprotectors and Pharmacology of the Institute of Bioorganic Chemistry named after A. S. Sadykov, Academy of Sciences of Uzbekistan, for their technical assistance.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors declare no conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
All animal experiments were conducted in accordance with the guidelines of the Ethics Committee of Andijan State University (protocol No. 07/2023). This research did not involve human participants.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Author Contributions
- Ibrohim R. Askarov: Project administration, Methodology.
- Yusupjon T. Isayev: Data curation, Formal analysis.
- Sanjar A. Rustamov: Investigation, Validation.
- Baxrom A. Mirzaabdullayev: Resources.
- Dostonbek U. Egamberdiyev: Software.
- Lazizbek Makhmudov: Supervision, Writing – review & editing.
- Dilfuza A. Amankulova: Data Collection, Analysis, Writing – review & editing.
References
- Askarov I.R. Sirli tabobat. Tashkent – Fan va texnologiyalar nashriyot-matba uyi – 2024, 1082 b.
- Isaev YT, Rustamov SA, Mirzaabdullayev BA. Determination of phenolic compound content and anti-inflammatory dose of the dietary supplement “PULMACLIN”. J Chem Goods Tradit Med. 2025;4(5):[in press]. doi:10.55475/jcgtm/vol4.iss5.2025.562
CrossRef - Askarov IR, Atakulova N. Assessment of the eco-biochemical composition of Raphanus sativus leaves grown in Uzbekistan. BIO Web Conf. 2025;194:01038. doi:10.1051/bioconf/202519401038
CrossRef - Frühbeck G, Gómez-Ambrosi J, Muruzábal FJ, Burrell MA. The adipocyte: a model for integration of endocrine and metabolic signaling in energy metabolism regulation. Am J Physiol Endocrinol Metab. 2001;280(6):E827-E847.
CrossRef - Stefanov AV, ed. Preclinical Investigation of Medicinal Substances (Methodological Guidelines). Kyiv; 2002:587.
- Mironov AN, ed. Guidelines for Preclinical Study of Non-steroidal Anti-inflammatory Drugs. In: Guidelines for Preclinical Drug Studies, Part 1. Moscow; 2012:767-776.
- Sernov LN, Gatsura VV. Elements of Experimental Pharmacology. Moscow; 2000:352.
- Askarov IR, Kodirov K. Helenium L. plant flower and leaf growing in Uzbekistan. BIO Web Conf. 2025;194:01037. doi:10.1051/bioconf/202519401037
CrossRef - Morris GM, Huey R, Lindstrom W, et al. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J Comput Chem. 2009;30(16):2785-2791.
CrossRef - Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking. J Comput Chem. 2010;31(2):455-461.
CrossRef - Isaev YT, Rustamov SA, Mirzaabdullayev BA. Determination of acute toxicity of the biologically active compound Pulmaklin. Xalq Tabobati Plus. 2026;1(26):[in press].
- Karin M. NF-κB as a critical link between inflammation and cancer. Cold Spring Harb Perspect Biol. 2009;1(5):a000141. doi:10.1101/cshperspect.a000141
CrossRef - Hotamisligil GS. Inflammation and metabolic disorders. 2006;444(7121):860-867. doi:10.1038/nature05485
CrossRef - Smith WL, DeWitt DL, Garavito RM. Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem. 2000;69:145-182. doi:10.1146/annurev.biochem.69.1.145
CrossRef - Kalgutkar AS, Crews BC, Marnett LJ. Kinetics of the interaction of nonsteroidal anti-inflammatory drugs with prostaglandin endoperoxide synthase-1 studied by limited proteolysis. 1996;35(28):9076-9082. doi:10.1021/bi9605752
CrossRef - Kishimoto T. Interleukin-6: discovery of a pleiotropic cytokine. Arthritis Res Ther. 2006;8(Suppl 2):S2. doi:10.1186/ar1916
CrossRef - Miller JF, Mitchell GF. Cell to cell interaction in the immune response. J Exp Med. 1968;128(4):801-820. doi:10.1084/jem.128.4.801
CrossRef - Davis RJ. Signal transduction by the JNK group of MAP kinases. 2000;103(2):239-252. doi:10.1016/s0092-8674(00)00116-1
CrossRef - Lu N, Malemud CJ. Extracellular Signal-Regulated Kinase: a regulator of cell growth, inflammation, chondrocyte and bone cell receptor-mediated gene expression. Int J Mol Sci. 2019;20(15):3792. doi:10.3390/ijms20153792
CrossRef - Karin M. NF-kappaB as a critical link between inflammation and cancer. Cold Spring Harb Perspect Biol. 2009;1(5):a000141. doi:10.1101/cshperspect.a000141
CrossRef - Sampaio ALF, et al. Biphasic Modulation of NOS Expression, Protein and Nitrite Products by Hydroxocobalamin Underlies Its Protective Effect in Endotoxemic Shock. Mediators Inflamm. 2013;2013:741804. doi:10.1155/2013/741804
CrossRef - Kishimoto T. Interleukin-6: From Basic Science to Medicine—40 Years in Immunology. Annu Rev Immunol. 2005;23:1-21. doi:10.1146/annurev.immunol.23.021704.115806
CrossRef
Abbreviations
AEA – Anti-exudative activity; BAC – Biologically active compound; COX-2 – Cyclooxygenase-2; IL-6 – Interleukin-6; MAPK – Mitogen-activated protein kinase; NF-κB – Nuclear factor kappa-B; TNF-α – Tumor necrosis factor-alpha.

