
Manuscript accepted on :09-06-2026
Published online on: 16-06-2026
Plagiarism Check: Yes
Reviewed by: Dr. Sunil dutt Shukla and Dr. Karthikeyan
Second Review by: Dr. Moumita Hazra and Dr. Anjaneyulu Vinukonda
Final Approval by: Dr H Fai Poon
Upik Rahmi1,2* , Hanna Goenawan2, Nova Sylviana2, Iwan Setiawan2, Hamidie Ronald DanielRay3, Uni Gamayani4,6, Yuni Susanti Pratiwi2, Achmad Adam5,6and Farida Murtiani7
, Hanna Goenawan2, Nova Sylviana2, Iwan Setiawan2, Hamidie Ronald DanielRay3, Uni Gamayani4,6, Yuni Susanti Pratiwi2, Achmad Adam5,6and Farida Murtiani7
1Nursing Program Study, Faculty of Sport and Health Education, UniversitasPendidikan Indonesia, Bandung, Indonesia.
2Department of Biomedical Science, Faculty of Medicine, UniversitasPadjadjaran, Bandung, Indonesia.
3Department of Medicine, Faculty of Medicine, Universitas Pendidikan Indonesia, Bandung, Indonesia.
4Department of Neurology, Faculty of Medicine, UniversitasPadjadjaran, Bandung, Indonesia.
5Department of Neurosurgery Faculty of Medicine UniversitasPadjadjaran, Bandung, Indonesia.
6Hasan Sadikin Hospital, Bandung, Indonesia.
7Department of Research, Sulianti Saroso Infectious Disease Hospital, Jakarta, Indonesia.
Corresponding Author E-mail: upikrahmi@upi.edu
Abstract
A sedentary lifestyle contributes to impaired hippocampal plasticity and may increase vulnerability to cognitive decline and neurodegenerative disorders. Physical exercise can regulate genes associated with synaptic function and neuroplasticity; however, the optimal exercise intensity for simultaneously modulating multiple neuroplasticity-related genes remains unclear. This study evaluated the effects of low-, moderate-, and high-intensity aerobic exercise on hippocampal mRNA expression of FNDC5/Irisin, EGR-1, p53, and Synaptophysin in male Wistar rats. Twenty rats were randomly allocated into four groups: sedentary control, low-intensity (10 m/min), moderate-intensity (20 m/min), and high-intensity (30 m/min) treadmill exercise. Interventions were performed for 8 weeks (5 sessions/week; 30 min/session). Gene expression was quantified using real-time PCR and analyzed using one-way ANOVA followed by LSD post hoc testing. Moderate-intensity exercise significantly increased hippocampal expression of FNDC5/Irisin (2.124 ± 0.448 vs 0.683 ± 0.202; p = 0.000), EGR-1 (1.158 ± 0.548 vs 0.605 ± 0.375; p = 0.032), p53 (3.031 ± 0.868 vs 1.018 ± 0.272; p = 0.010), and Synaptophysin (2.604 ± 1.310 vs 0.793 ± 0.548; p = 0.016) compared with sedentary controls. Low- and high-intensity exercise produced less consistent responses. The findings suggest that moderate-intensity aerobic exercise induces a more favorable molecular profile associated with neuroplasticity, potentially reflecting an optimal balance between neurotrophic signaling and the regulation of oxidative stress. This study provides comparative evidence supporting moderate-intensity exercise as a promising non-pharmacological strategy to promote hippocampal health and prevent cognitive decline.
Keywords
Aerobic Exercise; Gene Expression Regulation; Hippocampus; Neuroplasticity
| Copy the following to cite this article: Rahmi U, Goenawan H, Sylviana N, Setiawan I, Ray H. R. D, Gamayani U, Pratiwi Y. S, Adam A, Murtiani F. Moderate-Intensity Aerobic Exercise Enhances Hippocampal Neuroplasticity-Related Gene Expression: A Comparative Study in Rats. Biomed Pharmacol J 2026;19(2). |
| Copy the following to cite this URL: Rahmi U, Goenawan H, Sylviana N, Setiawan I, Ray H. R. D, Gamayani U, Pratiwi Y. S, Adam A, Murtiani F. Moderate-Intensity Aerobic Exercise Enhances Hippocampal Neuroplasticity-Related Gene Expression: A Comparative Study in Rats. Biomed Pharmacol J 2026;19(2). Available from: https://bit.ly/4uDF17w |
Introduction
A sedentary lifestyle, characterized by prolonged sitting or low physical activity,1 is a recognized risk factor for unhealthy brain aging, cognitive impairment, and neurodegenerative conditions, including Alzheimer’s disease. At the molecular level, these changes are accompanied by neurovascular dysfunction, reduced neurogenesis, and alterations in synaptic plasticity. Regular physical exercise is known to improve cognitive performance and brain health by promoting neurogenesis, angiogenesis, and synaptogenesis, as well as upregulating genes that support neuronal survival and plasticity.2
Among the key genes implicated in these processes are FNDC5/Irisin, EGR-1, p53, and Synaptophysin.Exercise-induced FNDC5/Irisin has been linked to enhanced synaptic plasticity and neurogenesis through neurotrophic pathways. EGR-1, an immediate-early gene, plays a pivotal role in memory consolidation. p53, traditionally known as a tumor suppressor, is also involved in neuronal homeostasis and synaptic modulation. Synaptophysin, a presynaptic vesicle protein, is widely used as a marker for synaptic density and functional connectivity. 3–5
The effects of physical exercise on the expression of one or two of these genes, often focusing on a single exercise intensity or using disease-specific animal models, have been explored in earlier research. For example, Wrann et al. demonstrated that aerobic exercise increases FNDC5/Irisin expression via the PGC-1α pathway,5. At the same time, Azimi et al. showed that moderate treadmill exercise ameliorates amyloid-β-induced cognitive deficits by upregulating FNDC5/Irisin and BDNF.6However, no study to date has systematically compared multiple exercise intensities while simultaneously evaluating the hippocampal expression of all four genesFNDC5/Irisin, EGR-1, p53, and Synaptophysinin the same experimental setting.
This integrated approach is essential for determining the optimal exercise intensity to enhance neuroplasticity-related gene expression, which may have translational value in designing targeted exercise interventions for cognitive health promotion and neurodegenerative disease prevention. Therefore, this study was conducted to evaluate the effects of three different levels of aerobic exercise intensity, namely i) low, ii) moderate, and iii) high,on the hippocampal mRNA expression of FNDC5/Irisin, EGR-1, p53, and Synaptophysin in male Wistar rats.
Materials and Methods
Animals and Ethical Approval
Male Wistar rats (Rattus norvegicus) aged 8–12 weeks and weighing 200–225 g were utilized in the present study. All animals were housed under controlled laboratory conditions, including a temperature of 22 ± 2 °C, relative humidity of 50–60%, and a 12-h light/dark cycle. Standard pellet feed and water were provided ad libitum throughout the experimental period.
Using G*Power software version 3.1, an a priori calculation was carried out with α = 0.05, statistical power of 0.80, and an effect size based on prior findings, indicating that a minimum of 8 rats per group was required.7
Due to facility constraints, n=5 animals per group were used, which is acknowledged as a limitation in statistical power.
Experimental Design and Grouping
Animals were randomly assigned to one of four groups based on aerobic exercise intensity measured in meters/minutes (m/min). The groups are: sedentary control (no exercise intervention) (Group 1), the low-intensity exercise group (Group 2) trained at a treadmill speed of 10 m/min, while the moderate- and high-intensity groups (Groups 3 and 4) exercised at 20 m/min and 30 m/min, respectively. Random allocation was conducted using a computer-generated sequence, and PCR analyses were performed by investigators blinded to the experimental grouping.
Determination of Exercise Intensity
Exercise intensity was determined based on blood lactate concentrations measured during a preliminary pilot study (n = 3 at each speed), obtained via tail vein sampling 5 min post-exercise. Lactate concentrations were as follows:i) low intensity (10 m/min): 2.5 ± 0.3 mmol/L, ii) Moderate intensity(20 m/min): 4.0 ± 0.4 mmol/L, and iii) high intensity(30 m/min): 6.8 ± 0.5 mmol/L. These values align with established thresholds for aerobic exercise intensities in rodents.6
Exercise Procedure
All exercise sessions were conducted between 08:00 and 10:00 AM to minimize circadian variation. Before the experimental period, animals underwent 2 weeks of treadmill habituation (5–10 m/min for 10 min/day). The exercise intervention was conducted for 8 weeks, 5 days per week, for 30 min per session, on a motorized treadmill (IDEAS, Indonesia).
A sham(Negative) control procedure was applied to the sedentary group by placing animals on the stationary treadmill for 5 min daily to control for handling and environmental exposure.
Tissue Collection
Twenty-four hours after the last exercise session, animals were anesthetized with 5% isoflurane in oxygen until respiration ceased.Following decapitation, the hippocampus was quickly dissected on ice, snap-frozen using liquid nitrogen, and stored at –80 °C for subsequent mRNA expression analysis. No lung tissue was collected for this study, thereby correcting previous reporting inconsistencies.
RNA Extraction and Real-Time PCR
Total RNA isolation from the hippocampus was carried out using TRIzol reagent (Bioline, UK) according to the manufacturer’s protocol. RNA purity and concentration were measured using spectrophotometry at 260/280 nm. cDNA synthesis and PCR amplification were performed using a One-Step RT-PCR Kit (Bioline, UK) on a real-time thermal cycler (Model one step MyTaq Kit).Real-time PCR was performed with an initial denaturation at 95 °C for 3 min, followed by 40 cycles comprising denaturation, annealing, and extension phases. GAPDH was utilized as the internal reference control. Primer sequences along with their corresponding annealing temperatures are presented in Table 1.
Table 1: Primers used in PCR
| Gene Symbol | Gene Bank | Primer Sequence | Product Length | Tm Annealing | GC% | Author (year) |
| FNDC5 | NM_001270981.2 | GATCATCGTCGTGGTCCTCTT | 116 | 59.59 | 52.38 | Park et al. (2020)1 |
| ATGCACTCTTGGTTTTTTCCTT | 56.97 | 36.36 | ||||
| EGR 1 | NM_012551.2 | CTCGCTCGGATGAGCTT ACA | 159 | 59.62 | 55.00 | Park et al. (2020)1 |
| TCCCACAAATGTCACAGGCA | 59.82 | 50.00 | ||||
| p53 | NM_030989.3 | TCGAGATGTTCCGAGAGCTGAATG | 107 | 62.51 | 50.00 | Wrann et al. (2013)5 |
| CTTCTTGGTCTTCGGGTAGCTG | 60.68 | 54.55 | ||||
| synaptophysin | NM_012664.3 | CGGAATACTTGGAGGCTGGG | 158 | 60.18 | 60.00 | Höttinget al. (2012)8 |
| ACAATACCGAAGGGCACAGG | 60.04 | 55.00 |
Quality control measures included: melting curve analysis to verify amplicon specificity, primer efficiency testing (acceptable range: 90–110%), and No-template controls (NTC) in each run to rule out contamination.
Statistical Analysis
Data are reported as mean ± SD. Statistical differences between groups were assessed using one-way analysis of variance (ANOVA) with LSD post hoc analysis, with significance set at p < 0.05. All statistical procedures were conducted using SPSS 26.0 (IBM Corp., USA).
Result
FNDC5/Irisin mRNA expressions in hippocampus tissue of male Wistar rats
Figure 1 shows the analysis of FNDC5/Irisin mRNA expression for all groups. There was a significant increase in FNDC5/irisin mRNA expression in the moderate-intensity exercise group (2.1243 ± 0.44793) compared to the control group (0.6829 ± 0.20249) (p = 0.000).
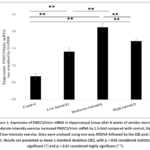 |
Figure 1: Expression of FNDC5/Irisin mRNA in hippocampal tissue after 8 weeks of aerobic exercise. Moderate-intensity exercise increased FNDC5/Irisin mRNA by 1.5-fold compared with control, high-, and low-intensity exercise. Data were analyzed using one-way ANOVA followed by the LSD post hoc test. |
EGR-1 mRNA expression in the hippocampus tissue of male Wistar rats
Figure 2 shows the analysis of EGR-1 mRNA expression for all groups. There was a significant increase in EGR-1 mRNA expression in the moderate-intensity exercise group (1.1576 ± 0.54760) compared to the control group (0.6046 ± 0.37535) (p = 0.032).
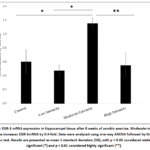 |
Figure 2: EGR-1 mRNA expression in hippocampal tissue after 8 weeks of aerobic exercise. Moderate-intensity exercise increases EGR-1mRNA by 0.4-fold. Data were analyzed using one-way ANOVA followed by the LSD post hoc test. |
p53 mRNA expressions in the hippocampus tissue of male Wistar rats
Figure 3 shows the analysis of p53 mRNA expression for all groups. There was a significant increase inp53 mRNA expression in the moderate-intensity exercise group (3.0306±0.86837) compared to the control group (1.0175 ± 0.27191) (p = 0.01).
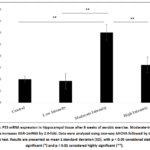 |
Figure 3: P53 mRNA expression in hippocampal tissue after 8 weeks of aerobic exercise. Moderate-intensity exercise increases EGR-1mRNA by 2.0-fold. Data were analyzed using one-way ANOVA followed by the LSD post hoc test. |
Synaptophysin mRNA expression in the hippocampus tissue of male Wistar rats
Figure 4 shows the analysis of Synaptophysin mRNA expression for all groups. A significant elevation in Synaptophysin mRNA expression was observed in the moderate-intensity exercise group(2.6038 ± 1.31006) compared to the control group (0.7925 ± 0.54764) (p=0.016).
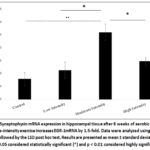 |
Figure 4: Synaptophysin mRNA expression in hippocampal tissue after 8 weeks of aerobic exercise. Moderate-intensity exercise increases EGR-1mRNA by 1.5-fold. Data were analyzed using one-way ANOVA followed by the LSD post hoc test. |
Discussion
This study represents the first systematic comparison of the effects of three distinct intensities of aerobic exercise on the simultaneous hippocampal expression of four neuroplasticity-related genes,namely i) FNDC5/Irisin, ii) EGR-1, iii) p53, and iv) Synaptophysinin male Wistar rats. The results demonstrate that moderate-intensity exercise produces the most consistent and significant upregulation across all four genes, suggesting an optimal balance between neurotrophic stimulation and oxidative stress regulation.
Our findings showed around a 3.1-fold increase in FNDC5/Irisin mRNA expression in the moderate-intensity exercise group compared with the sedentary control group, exceeding the fold changes observed in both low- and high-intensity groups. FNDC5/Irisin is cleaved to produce irisin, a myokine that promotes BDNF release via the PGC-1α–FNDC5–BDNF pathway, enhancing synaptic plasticity and neurogenesis.5Previous findings from Wrann et al. and Azimi et al. reported FNDC5/Irisin upregulation with exercise.However, these studies either used a single intensity or focused on disease models.5,6 Our simultaneous multi-intensity design suggests that moderate-intensity exercise achieves the optimal physiological window for activating this pathway without inducing excessive oxidative stress.
In this study, EGR-1, an immediate-early gene, increased by approximately1.9-fold with moderate-intensity exercise. This transcription factor regulates downstream genes involved in memory consolidation, LTP maintenance, and synaptic remodeling.9 While low-intensity exercise elicited minimal changes, and high-intensity exercise increased variability, our findings align with Bozon et al.,who showed that EGR-1 induction is tightly linked to stimulus intensity.10,11Importantly, the concurrent upregulation of EGR-1 and FNDC5/Irisin in our study suggests a possible synergistic effect, where EGR-1 primes neuronal circuits while FNDC5/Irisin supports structural remodeling.
Although best known as a tumor suppressor, p53 plays a functionally dual role in neurons that must be distinguished carefully: a pro-apoptotic role under conditions of severe or irreparable DNA damage, and a neuroprotective, pro-plasticity role in postmitotic neurons under sublethal cellular stress.¹² We observed approximately a 2.98-fold increase in p53 mRNA expression in the moderate-intensity exercise group. In the context of repetitive moderate aerobic exercise, this upregulation is more consistent with the neuroprotective arm of p53 signaling. Merlo et al. demonstrated through whole-genome ChIP analysis that p53 directly regulates synaptic function genes in postmitotic neurons and confers protection against tau-mediated neurodegeneration, with this regulatory function conserved in mammals11. Furthermore, Rahmi et al., in a recent narrative review, confirmed that LTP induction in hippocampal neurons correlates with increased p53 transcription and that p53 interacts with downstream mediators, including immediate early genes such as EGR-1, to regulate synaptic remodeling12. Together, these findings suggest that the moderate exercise-induced increase in p53 observed here likely reflects an adaptive, plasticity-promoting response rather than a pro-apoptotic signal.
It must be acknowledged, however, that the present study measured only p53 mRNA expression and not its downstream effectors. Without concurrent measurement of p21 (a marker of cell cycle arrest and senescence), Bcl-2/Bax ratio, or caspase-3 activation, definitive attribution of the p53 signal to plasticity-associated rather than stress-associated pathways remains an interpretive limitation. Future studies should incorporate these markers to disambiguate the functional role of p53 upregulation following exercise. In our study, high-intensity exercise resulted in lower p53 expression than moderate-intensity exercise. This may reflect active suppression of the neuroprotective p53 axis under excessive oxidative stress, consistent with the observation that elevated ROS at high exercise intensities can disrupt DNA damage-sensing pathways, ultimately impairing rather than enhancing synaptic homeostasis.13
Findings from our study showed that Synaptophysin expression increased about2.6-fold in the moderate-intensity group, suggesting enhanced presynaptic vesicle density and neurotransmission efficiency. This aligns with Vaynman et al, who reported increased synaptophysin levels following exercise-induced synaptic plasticity.14The parallel increase in synaptophysin and FNDC5/Irisin implies that molecular signaling from muscle to brain can translate into structural synaptic changes.
The inconsistent and attenuated molecular responses observed in the high-intensity exercise group across all four genes warrant deeper mechanistic investigation beyond a single oxidative-stress hypothesis. Several non-mutually exclusive explanations may account for this pattern.
First, the blood lactate concentration in the high-intensity group (6.8 ± 0.5 mmol/L) suggests that exercise at 30 m/min likely approached or exceeded the anaerobic threshold in Wistar rats, a condition associated not only with increased ROS production but also with elevated pro-inflammatory cytokine expression. Jahangiri et al. demonstrated that overtraining in rats significantly increased hippocampal TNF-α and IL-1β levels and led to impaired spatial learning and memory, implicating neuroinflammatory activation as a mechanism by which excessive exercise intensity disrupts hippocampal molecular homeostasis15. Second, high-intensity exercise is associated with elevated circulating corticosterone, a glucocorticoid that has been shown to suppress hippocampal neuroplasticity by attenuating AMPK-dependent signaling and inducing dendritic retraction in CA1 neurons16. This hormonal mechanism may compound the direct oxidative effects on gene transcription. Third, the observed high intragroup variability in gene expression among the high-intensity group — reflected by the relatively large standard deviations — may indicate that individual animals responded heterogeneously, possibly due to differences in fitness level, stress tolerance, or recovery capacity, given the fixed-intensity protocol. Lee et al. confirmed in spontaneously hypertensive rats that the beneficial effects of exercise on hippocampal synaptic density and spatial memory were abolished at high intensity, with a concomitant increase in hippocampal free radical levels, suggesting that the oxidative threshold for neuroplasticity impairment may be relatively narrow and model-dependent17.
Collectively, our data indicate that moderate-intensity exercise optimally activates a molecular profile conducive to neuroplasticity: i) FNDC5/Irisin for neurotrophin release, ii) EGR-1 for transcriptional priming, iii) p53 for synaptic regulation, and iv) synaptophysin for structural connectivity. We hypothesize that moderate intensity maintains ROS production within an adaptive range, allowing activation of AMPK–PGC-1α pathways without triggering NF-κB–mediated inflammatory cascades. In contrast, high-intensity exercise may exceed this oxidative threshold,impairmolecular signaling, and reduce net neuroplasticity benefits.15
Our findings expand upon previous work by integrating four genes into a single experimental framework and intensity comparison. For instance, Wrann et al. examined FNDC5/Irisin alone,5 while Azimi et al. explored FNDC5/Irisin and BDNF in an Alzheimer’s model.6 To our knowledge, no prior study has directly contrasted low-, moderate-, and high-intensity exercise for these four molecular targets simultaneously in healthy hippocampal tissue.
These results suggest that moderate-intensity aerobic exercise may be an effective non-pharmacological strategy to promote hippocampal synaptic health and prevent cognitive decline. The molecular profile identified here provides a potential biomarker framework for tailoring exercise prescriptions in both healthy and at-risk populations, such as older adults or individuals with early cognitive impairment.
This study has several limitations. First, the relatively small sample size (n = 5 per group) may have reduced the statistical power of the analysis and led to an underestimation of true effects. Second, the exclusive use of male rats leaves potential sex-based differences in neuroplasticity unexplored. A key limitation is the lack of direct measurements of key mechanistic components, such as oxidative stress, BDNF, and neuroinflammatory markers, which prevents a definitive mechanistic interpretation. Consequently, while the proposed model is supported by existing literature, the causal links between changes in gene expression and reduced oxidative damage remain hypothetical. Furthermore, the observed molecular changes in the hippocampus were not correlated with functional outcomes, as no behavioral or cognitive tests were conducted. Finally, the focus solely on the hippocampus limits the findings, as other brain regions involved in cognition were not examined. Future studies should therefore incorporate these biochemical measurements and behavioral assays to directly link molecular changes to cognitive function.
Conclusions
Among the three exercise intensities examined, moderate-intensity treadmill exercise elicited the most consistent and significant upregulation of all four neuroplasticity-related genes — FNDC5/Irisin, EGR-1, p53, and Synaptophysin — in the hippocampus of male Wistar rats, suggesting an optimal balance between neurotrophic stimulation and oxidative stress regulation at the molecular level. These findings provide preliminary molecular evidence that moderate-intensity aerobic exercise may favourably modulate hippocampal gene expression profiles associated with synaptic plasticity. However, several important caveats must be noted before any translational inference can be made. The present study was conducted exclusively in a rodent model with a small sample size (n = 5 per group) and did not directly measure protein expression, oxidative stress markers, neuroinflammatory mediators, or cognitive behavioural outcomes. Accordingly, the extent to which these mRNA-level changes translate into functional neuroplasticity — and, critically, whether equivalent molecular responses would be observed in humans — remains to be established. Species differences in exercise physiology, brain metabolism, and hippocampal gene regulation limit the direct extrapolation of these findings to clinical or public health contexts.
Within these limitations, this work extends beyond single-gene or single-intensity studies by integrating four molecular markers within the same experimental framework. The multi-gene, multi-intensity design offers a useful preclinical reference for hypothesis generation regarding exercise dose–response relationships in neuroplasticity research. Future studies incorporating larger sample sizes, both sexes, protein-level and behavioural endpoints, direct measurements of oxidative stress and neurotrophins, and ultimately translational work in human cohorts will be necessary to determine whether moderate-intensity exercise can serve as an evidence-based, non-pharmacological strategy for hippocampal health promotion and cognitive decline prevention in at-risk populations.
Acknowledgement
We want to extend our sincere gratitude to the entire research team of the Department of Nursing, Faculty of Sport and Health Education, Universitas Pendidikan Indonesia. We would like to thank the Faculty of Medicine, UniversitasPadjadjaran, and the supporting team for their support of this study from the outset.
Funding Sources
The study was supported by grant funding from LembagaPengelola Dana Pendidikan from the Indonesian Ministry of Finance (Number:0001454/TRP/D/PDD-2020).
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
All procedures were conducted in accordance with the ARRIVE guidelines and approved by the Padjadjaran University Research Ethics Committee (Approval No. 369/UN6.C.6.30/PT.01.04/2022).
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Author Contributions
- Upik Rahmi: Conceptualization, Methodology, Writing – Original Draft, Writing – Review & Editing
- Hanna Goenawan: Conceptualization, Methodology, Supervision, Writing – Original Draft
- Nova Sylviana: Conceptualization, Methodology, Writing – Original Draft
- Iwan Setiawan: Conceptualization, Methodology, Writing – Original Draft
- Hamidie Ronald DanielRay: Data Collection, Writing – Original Draft, Writing – Review & Editing
- UniGamayani: Data Collection, Supervision, Writing – Original Draft
- Yuni Susanti Pratiwi: Project Administration
- Achmad Adam: Funding Acquisition, Resources, Writing – Original Draft
- Farida Murtiani: Visualization, Analysis, Writing – Review & Editing
Refernces
- Park JH, Moon JH, Kim HJ, Kong MH, Oh YH. Sedentary Lifestyle: Overview of Updated Evidence of Potential Health Risks. Korean J Fam Med. 2020 Nov;41(6):365–73. doi:10.4082/kjfm.20.0165 PubMed PMID: 33242381.
CrossRef - Erickson KI, Hillman C, Stillman CM, Ballard RM, Bloodgood B, Conroy DE, et al. Physical Activity, Cognition, and Brain Outcomes: A Review of the 2018 Physical Activity Guidelines. Med Sci Sports Exerc. 2019 Jun 1;51(6):1242–51. doi:10.1249/MSS.0000000000001936 PubMed PMID: 31095081.
CrossRef - Zhao Y, Jia M, Chen W, Liu Z. The neuroprotective effects of intermittent fasting on brain aging andneurodegenerative diseases via regulating mitochondrial function. Free RadicBiol Med. 2022 Mar;182:206–18. doi:10.1016/j.freeradbiomed.2022.02.021 PubMed PMID: 35218914.
CrossRef - Hötting K, Röder B. Beneficial effects of physical exercise on neuroplasticity and cognition. NeurosciBiobehav Rev. 2013 Nov;37(9 Pt B):2243–57. doi:10.1016/j.neubiorev.2013.04.005 PubMed PMID: 23623982.
CrossRef - Wrann CD, White JP, Salogiannnis J, Laznik-Bogoslavski D, Wu J, Ma D, et al. Exercise Induces Hippocampal BDNF through a PGC-1α/FNDC5 Pathway. Cell Metab. 2013 Nov 5;18(5):649–59. doi:10.1016/J.CMET.2013.09.008 PubMed PMID: 24120943.
CrossRef - Azimi M, Gharakhanlou R, Naghdi N, Khodadadi D, Heysieattalab S. Moderate treadmill exercise ameliorates amyloid-β-induced learning and memoryimpairment, possibly via increasing AMPK activity and up-regulation of the PGC-1α/FNDC5/BDNF pathway. Peptides (NY). 2018 Apr;102:78–88. doi:10.1016/j.peptides.2017.12.027 PubMed PMID: 29309801.
CrossRef - Faul F, Erdfelder E, Lang AG, Buchner A. G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav Res Methods. 2007;39(2):175–91. doi:10.3758/BF03193146 PubMed PMID: 17695343.
CrossRef - Hötting K, Reich B, Holzschneider K, Kauschke K, Schmidt T, Reer R, et al. Differential cognitive effects of cycling versus stretching/coordination trainingin middle-aged adults. Health Psychol. 2012 Mar;31(2):145–55. doi:10.1037/a0025371 PubMed PMID: 21895371.
CrossRef - Guzowski JF, McNaughton BL, Barnes CA, Worley PF. Environment-specific expression of the immediate-early gene Arc in hippocampal neuronal ensembles. Nat Neurosci. 1999 Dec;2(12):1120–4. doi:10.1038/16046 PubMed PMID: 10570490.
CrossRef - Bozon B, Davis S, Laroche S. Regulated transcription of the immediate-early gene Zif268: mechanisms and genedosage-dependent function in synaptic plasticity and memory formation. Hippocampus. 2002;12(5):570–7. doi:10.1002/hipo.10100 PubMed PMID: 12440572.
CrossRef - Jacobson BH, Smith D, Fronterhouse J, Kline C, Boolani A. Assessment of the benefit of powered exercises for muscular endurance and functional capacity in elderly participants. J Phys Act Health. 2012;9(7):1030–5. doi:10.1123/JPAH.9.7.1030 PubMed PMID: 22971882.
CrossRef - Vousden KH, Lu X. Live or let die: the cell’s response to p53. Nat Rev Cancer. 2002 Aug;2(8):594–604. doi:10.1038/NRC864 PubMed PMID: 12154352.
CrossRef - Vaynman S, Gomez-Pinilla F. License to run: exercise impacts functional plasticity in the intact and injured central nervous system by using neurotrophins. Neurorehabil Neural Repair. 2005 Dec;19(4):283–95. doi:10.1177/1545968305280753 PubMed PMID: 16263961.
CrossRef - Radak Z, Chung HY, Koltai E, Taylor AW, Goto S. Exercise, oxidative stress and hormesis. Aging Res Rev. 2008 Jan;7(1):34–42. doi:10.1016/J.ARR.2007.04.004 PubMed PMID: 17869589.
CrossRef

