
Manuscript accepted on :20-05-2026
Published online on: 10-06-2026
Plagiarism Check: Yes
Reviewed by: Dr. Mandal, Amritlal
Second Review by: Dr. Moumita Hazra Drmoumita
Final Approval by: Dr. Ian James Martin
Rinad Sleman Aldhali1, Medhat Rehan2 and Imen Ben Abdelmalek1*
and Imen Ben Abdelmalek1*
1Biology Department, College of Science, Qassim University, Burydah, Saudi Arabia,
2Department of Plant Production, College of Agriculture and Food, Qassim University, Buraydah, Saudi Arabia
Corresponding Author E-mail: mm.abdulmalek@qu.edu.sa
Abstract
Streptomyces tricolor HM10 was investigated for extracellular protease and protease inhibitor production, partial purification, biochemical characterization, and gene-level confirmation. ISP-4 medium was the most suitable for production, with maximum protease inhibitor activity observed after 2 days of incubation. Casein significantly enhanced enzyme production, indicating its role as an inducer. Gel filtration chromatography on Sephacryl S-200 successfully separated protease and inhibitor activities, confirming their distinct molecular nature. The protease showed optimal activity at pH 9.0 and 50 °C, while the inhibitor exhibited maximum activity at neutral pH and 60 °C. Both biomolecules demonstrated broad pH and thermal stability. Metal ions such as Mn²⁺ and Co²⁺ enhanced activity, whereas Hg²⁺ strongly inhibited both proteins. PMSF sensitivity confirmed the serine nature of the protease. The enzyme retained stability in the presence of non-ionic surfactants, suggesting potential industrial applicability. Additionally, strong antifungal activity was observed against plant pathogens, particularly Fusarium oxysporum. PCR amplification confirmed the presence of genes encoding serine protease, subtilase-type inhibitor, and CPBP metalloprotease, while one inhibitor gene was not detected. Overall, S. tricolor HM10 represents a promising source of thermostable alkaline protease and bioactive inhibitor with potential applications in biotechnology and biocontrol.
Keywords
Characterization; Enzyme inhibitor; Gene amplification; Serine proteinase; Streptomyces
| Copy the following to cite this article: Aldhali R. S, Rehan M, Abdelmalek I. B. Screening, Partial Purification, Biochemical and Molecular Characterization of Streptomyces tricolor HM10 Serine Protease and Protease Inhibitors. Biomed Pharmacol J 2026;19(2). |
| Copy the following to cite this URL: Aldhali R. S, Rehan M, Abdelmalek I. B. Screening, Partial Purification, Biochemical and Molecular Characterization of Streptomyces tricolor HM10 Serine Protease and Protease Inhibitors. Biomed Pharmacol J 2026;19(2). Available from: https://bit.ly/4uKJMwh |
Introduction
Proteases, also referred to as proteinases or proteolytic enzymes, are essential components of cell health and viability. They are involved in a variety of processes, including transcription, cell proliferation, differentiation, extracellular matrix remodelling, hormone processing, and the processing of biologically active peptides.1The hydrolysis of peptide bonds in proteins and polypeptides is catalysed by a broad class of enzymes known as proteases. Substrate specificity, active site, catalytic mechanism, pH, temperature optima, and stability profile are some of the characteristics that set them apart.There are several schemes for classifying proteases, which provide a wealth of relevant information about each protease. According to the Enzyme Commission (EC) classification, proteases belong to hydrolases (group 3), which hydrolyze peptide bonds (sub-group 4).2 Also, Proteases are regarded as the most beneficial and potent enzymes and represent 6% of the total proteins in the human genome and represent 1-5% of microbial genomes (such as bacteria, fungi, and viruses).3, 4
Protease enzymes play metabolic and regulatory functions, are basic in many biological processes, and offer a wide range in physiological, biochemical processes and in biotechnological applications.5, 6
Continuous neutrophil infiltration and activation frequently contribute to chronic inflammation. The current understanding is that neutrophil serine proteases play a major role in these disorders by causing proteolytic tissue destruction following their release from activated or dead neutrophils.6 In order to prevent the breakdown of connective tissue proteins including elastin, collagen, and proteoglycans, proteolytic activity appears to be strictly controlled in the extracellular and pericellular area. Serine proteases’ proteolytic activity is ultimately regulated by protein inhibitors that are members of three major families: serpins, chelonianins, and macroglobulins.7
Protease inhibitors are a widely used as class of antivirals, and these selective inhibitors can be used as a powerful tool to inactivate target proteases,8 so they are widely used in the treatment of diseases, such as influenza, HIV, malaria, cancer, and even degenerative diseases, such as Alzheimer’s disease and their treatment for HIV/AIDS and hepatitis C.9
Therapeutic inhibitors have been studied as a treatment for a number of viral and chronic inflammatory illnesses by controlling undesired proteolysis at inflammation sites. Corticosteroid medications are the mainstay of treatment. Corticosteroids, like any medications, can have unfavourable side effects, particularly when taken in high dosages for extended periods of time. Hyperglycemia, infection, gastritis, gastrointestinal bleeding, gastrointestinal ulceration or perforation, proximal myopathy, severe peripheral oedema, and psychiatric episodes are examples of corticosteroid toxicity.
In order to avoid these secondary effects, several reports are focused on the screening and the identification of new serine protease and inhibitors proteasewith no major side effects from different natural sources.
Actinomycetes, especially Streptomyces species, are a class of bacteria that are extensively used for the manufacture of commercially valuable secondary metabolites and enzymes for use in agriculture and medicine.10 Actinomycetes are present in soil and in many kinds of natural resources.11 Streptomyces is the most important genus for both industrial and human health care since it appears to produce a greater range of novel bio-functional molecules than any other genus.12 Nowadays, they generate a lot of secondary metabolites for pharmaceutical companies, which results in extensive research being done to find new serine proteases and protease inhibitors.
Streptomyces albogriseolus S-3253 is the original source of the proteinaceous protease inhibitor known as Streptomyces subtilisin inhibitor (SSI).13 Alkaline serine proteases are strongly inhibited by SSI, a dimer made up of two identical subunits. A wide range of Streptomyces species and their related species produce SSI and SSI-like protease inhibitors (SIL), which together make up the vast SSI family.14The widespread presence of SSI and SIL in actinobacteria points to a significant shared function for these inhibitors in the producers.15
The objective of our study is the screening,the production, biochemical,and molecular characterization of a novel protease and protease inhibitor of Streptomyces HM10 strain newly isolated from Qassim University soil.
Materials and Methods
Microorganisms and used media
Streptomyces tricolor HM10used in this study was isolated by Dr. Medhat Rehan (Department of Plant Production,College of Agriculture and Food) from the soil of Qassim University. The strainwas maintained by periodic transfer to fresh stock culture media every month and storedat 10 ºC.15Three culture media were prepared and tested, theSMF medium (calcium carbonate 0.2g, peptone 0.15g, glucose 0.9g in 100 mL distilled water), ISP-4 medium (glucose 1 g, potassium dihydrogen phosphate 0.1 g, magnesium sulfate 0.1 g, sodium chloride 0.1 g, ammonium sulfate 0.2 g, calcium carbonate 0.2 g, trace salts solution, in 100 mL distilled water, (pH 7.0) and the King’s Medium B Base (proteose peptone 0.2 g, dipotassium hydrogen phosphate 0.015g, magnesium sulphate heptahydrate 0.015g in 100 mL distilled water(pH 7.0).
For each culture medium, the strain was cultivated in media with or without 2% of casein as unique carbon source. Autoclaving for 20 min was then carried out at a temperature of 120ºC.
Culture conditions
A single colony of Streptomyces tricolor HM10 was used to inoculate a seed culture medium from King’s medium B for 24 h at 30ºC and 170 rpm on incubator shaker (Thermo Fisher Scientific) for agitation and aeration. One milliliterof the seed culture was used to inoculate each of the three media (100 mL).After that, the cultures were incubated for 72 hours at 30 °C with agitation (150 rpm). Following the incubation period, the cell pelletmass was discarded and the cell-free extract was recovered by centrifugation at10,000 rpm for 15 minutes.15
Partial purification
In accordance with Kalchev et al. (2013),15 partial purification was carried out. Samples were first centrifuged at 12,000 rpm for 40 minutes at 4°C to remove cell debris. The resulting supernatants were precipitated with 80% (NH₄)₂SO₄ at 4°C for two days, and the precipitated proteins were collected by centrifugation under the same conditions. The Bradford16 test was used to measure the protein content, and protease and protease inhibitor activities were measured using their respective protocols.
The precipitated proteins were resuspended in 20 mM Tris-HCl (pH 7.0) and dialyzed against the same buffer for two days to remove excess salts. A Sephacryl S-200 column (89 cm × 2 cm) that had been pre-equilibrated with 20 mM Tris-HCl, pH 7.0, containing 0.15 M NaCl was then filled with the dialysed samples. The same buffer was used to elute the proteins, and 3 mL fractions were gathered at a flow rate of 0.5 mL/min. Spectrophotometric monitoring of protein elution was conducted at 280 nm. Pooled fractions with protease or protease inhibitor activity were kept at 4°C for further molecular and biochemical investigation.
Protease assay activity
The protease activity in the culture supernatant was measured using the Tsuchida et al. technique17 with casein as a substrate. The enzyme activity was measured by incubating 200 µL of the crude extract with 500 µL of 1% (w/v) casein sodium salt (Sigma) in 100 mM buffer (pH 7) for 30 minutes at 37 °C. After stopping the reaction with 250 µL (0.44 M) of trichloroacetic acid (TCA), the mixture was left at room temperature for 15 minutes. After that, the reaction mixture was centrifuged for 15 minutes at 10,000 rpm to extract the non-reacted casein.18 A spectrophotometer (Agilent Technologies) was used to measure the activity at 280 nm.
Protease inhibitor assay
Protease inhibitor activities against trypsin were determined according to Tsuchida et al.17In brief, 200 µLof trypsin (1000 units/mg) was combined with 200 µL aliquot of Streptomyces sp. crude extract, and the mixture was pre-incubated for 15 minutes at room temperature. After that, 400 µL of 1% casein was added and the mixture was left at 37 °C for 30 minutes. The process was finally stopped by adding 250µL of 0.44M Trichloroacetic acid (TCA) solution. The absorbance at 280 nm was measured following centrifugation of the reaction mixture at 12,000 rpm for 15 minutes.Under standard test conditions, one unit of protease inhibitor activity (PIU) was defined as the decrease of one unit of absorbance at 280 nm of the TCA soluble casein hydrolysis product released by trypsin per minute.18 For comparative purposes, the inhibition percentage of the protease inhibitor activity was also calculated by comparing it to a control experiment.
Protein determination
The Bradford method16 was used to measure the protein concentration, using bovine serum albumin as a reference.
Effect of pH and temperature on the protease and its inhibitor activity and stability
The optimum pH of S. tricolorHM10 protease and its inhibitor activity were studied by varying pH values (3-13) at 37°C. The corresponding pH buffers, 100 mM sodium acetate buffer (pH 3-5), 100 mM potassium phosphate buffer (pH 6.0-7.0), 100 mM Tris-HCl buffer (pH 8.0-9,0), and 100 mM glycine-NaOH buffer (pH 10-13), were used to create the 1% casein substrate. The pH stability was assessed by using the same buffers to incubate the enzyme at different pH values between 3 and 13 for 12 hours at 4 °C. After centrifugation (30 minutes at 12,000 rpm), the remaining protease activity was measured using the conventional test procedure. Every measurement was made three times.
To analyze the effect of temperature, the activity of protease and its inhibitor were tested at different temperatures (4-100°C) for 30 min at the optimum selected pH of 9 and 7, respectively from the previous experiment. The thermal stability of the activities was evaluated by incubating the protease and its inhibitor at different temperatures (4-100°C) for 30 min in phosphate buffer (pH7.0) and glycine-NaOH buffer (pH 9.0). The above-described enzyme assay was used to quantify residual protease. Prior to incubation, the enzyme was thought to be 100% active.
Storage stability
Protease was stored in phosphate buffer at 4°C, and protease/inhibitor activity and protein concentration were monitored every 7 days.
Effect of metal ions and inhibitors on protease and its inhibitor activity
CaCl2, MnCl2, ZnSO4, CoSO4, CuSO4, NaCl, KCl, FeSO4, and MgSO4 were used to examine the impact of different metal ions (5 mM) on enzyme activity. EDTA, iodoacetate, and PMSF were used to investigate their effect on the protease and its inhibitor activity. The remaining enzyme and inhibitor activity were evaluated after their preincubation with metals and chelators for 30 minutes at 37°C.
Antifungal activity of purified protease as a biocontrol agent against some plant pathogenic fungi
The alkaline protease produced by Streptomyces tricolor HM10 was assessed for antifungal activity using the agar well diffusion method. Potato dextrose agar (PDA) plates were prepared and inoculated with fungal pathogens (Fusarium oxysporum, Sclerotinia sclerotiorum, Aspergillus niger, Rhizoctonia solani, Trichoderma spp., and Botrytis cinerea) by evenly spreading fungal spores or mycelial fragments onto the agar surface.
Wells of approximately 6 mm diameter were aseptically punched into the agar plates. Two different volumes (25 μL and 50 μL) of the crude alkaline protease enzyme solution were added separately into the wells. Plates were then incubated at 28 ± 2 °C for 5-7 days, depending on the fungal growth rate.
Toevaluate the protease’s antifungal activity, the diameter of the inhibitory zones surrounding each well was determined in millimetres following incubation. Control wells containing sterile buffer or enzyme-free medium were included to confirm the specificity of inhibition. All experiments were performed in triplicate, and mean values were reported.
Trypsin Inhibition Assay Using Gelatin Agar Plates
A gelatin agar plate protocol adapted from common protease diffusion techniques was used to measure trypsin inhibitory activity19. In brief, gelatin (1% w/v) was dissolved in agar (2% w/v) using the proper buffer (such as phosphate buffer, pH 7) to create agar plates. After sterilisation, the medium was transferred into Petri dishes and left to harden.
A sterile cork borer was used to punch uniformly sized wells into the gelatin agar. Trypsin solutions of varying trypsin (5, 10, 15, and 20 U) were prepared, and equal volumes were added either alone (trypsin control) or in combination with the inhibitor at two concentrations (50 and 150 mg/L). Plates were incubated at 37 °C for 18-24 hours to allow enzymatic digestion of gelatin.Following incubation, the plates were flooded with Bromophenol blue solution to stain the intact protein matrix. Excess stain was removed by rinsing with distilled water, revealing clear zones corresponding to areas of gelatin hydrolysis. The diameter of the clear zones was measured in millimeters, and inhibition was assessed by comparing the zones in the presence and absence of inhibitor
DNA extraction andStreptomyces HM10 gene amplification by PCR
ISP-4’smedium was used to maintain Streptomyces tricolor HM10. The bacterial culture was grown aerobically at pH 7.3, 30°C for 24 h for DNA isolation.
Genomic DNA was extracted under a UV-sterilized laminar flow hood with dedicated equipment using QIAamp DNA Micro kits (Qiagen) following the manufacturer’s protocol. Primers were designed according to the known DNA sequence of Streptomyces strain (Streptomyce citricolor212427). Usedprimer sequences are indicated in supplementary
Table 1.
The reaction mixture included a reaction buffer, 1.5 mM MgCl2, 0.2 mM dNTPs, 0.5 U GoTaq DNA polymerase (Promega), 0.4 mM of each primer, and 20 ng of genomic DNA. The PCR was conducted in a Thermocycler (BioRad) for thirty cycles. The following PCR cycling settings were employed: 30 cycles of 95°C for 30 seconds, 58°C for 30 seconds, and 72°C for 1 minute were followed by a two-minute initial denaturation at 94°C. For ten minutes, the last extension was at 72°C. On an agarose gel, the PCR-amplified genes were revealed.
Table 1: Forward and Reverse primer sequences used in the amplification of S. tricolor HM10 protease and protease inhibitor genes.
| GC (%) | Tm (°C) | Length (bp) | Sequence (5′–3′) | Primers | Gene |
| 52.6
50 |
62
62 |
19
20 |
GAGTTGACCATGCCGATCA
CGGGTTCTTCTCGTTCGATT |
F
R |
Serine protease |
| 60
61.1 |
62
62 |
20
20 |
TACAGGCAGACGAAGACCA
CCTCGGCTTCAACCTCAC |
F
R |
CPBP family metalloprotease |
| 55.6
55.6 |
61
61 |
18
18 |
GGCCGTGCTGAAGTACC
GTGATCGTGAAGACGTAGTACC |
F
R |
Protease inhibitor I42 family protein |
| 55.6
55.6 |
61
61 |
18
18 |
TGATCACCTCACCGTCGT
CGATCTCGCACCCGTTG |
F
R |
Subtilase-type protease inhibitor |
Agarose Gel Electrophoresis Analysis
Agarose gel electrophoresis was used to analyse the PCR products. In brief, agarose was dissolved in 1× TAE buffer and heated until fully melted to create 1% (w/v) agarose gel. The gel was poured into a casting tray with a comb to create wells after cooling to about 50–55 °C and adding 0.5 µg/mL of ethidium bromide (or a safe DNA dye like Gel Red).
Each PCR product (5 µL) was combined with loading dye and added to the wells along with a 100 bp DNA ladder. Electrophoresis was performed in 1× TAE buffer for 45–60 minutes at 90-110 V. A gel documentation system was then used to see DNA bands under UV light.
Statistical Analysis
All experiments were performed in triplicate, and results are expressed as mean ± standard deviation (SD). Statistical analysis was carried out using Microsoft Excel 2016or GraphPad Prism version 9 (GraphPad Software, San Diego, CA, USA). Differences between means were evaluated using one-way analysis of variance (ANOVA), and values of p< 0.05 were considered statistically significant.
Results
In this study, an extracellular alkaline protease and its inhibitor from Streptomyces tricolor HM10 were partially purified and characterised with interesting medicinal and biotechnology implications.
Streptomyces tricolor HM10 Protease and Inhibitor Production
Three liquid culture media (SMF, ISP-4, and King’s Medium B) were evaluated for the production of extracellular protease inhibitors by Streptomyces tricolor HM10. Based on these results, ISP-4 was found to be the most suitable forthe protease inhibitorproduction. The fermentation time course for protease inhibitor production by Streptomyces tricolor HM10 (Fig.1) showed that maximum activity was obtained after 2 daysof cultivation. Thereafter, a slight decline in activity was observed.
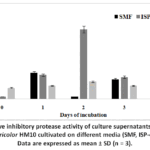 |
Figure 1: Relative inhibitory protease activity of culture supernatants obtained fromStreptomyces tricolor HM10 cultivated on different media (SMF, ISP-4 and King’s B). |
Protease production was further evaluated in ISP-4 medium in the presence and absence of casein as the unique nitrogen source. The effect of 2% casein on enzyme production was monitored over different incubation periods. As shown in Fig. 2, protease activity increased progressively with incubation time, reaching a maximum on the 9th day, followed by a gradual decline. The presence of casein significantly enhanced enzyme production at all time points, with peak activity (~100%) observed under Casein+ conditions compared to lower levels in its absence. These findings suggest that casein acts as an inducer, stimulating protease synthesis.
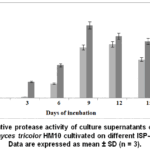 |
Figure 2: Relative protease activity of culture supernatants obtained from Streptomyces tricolor HM10 cultivated on different ISP-4 media. |
Partial purification of Streptomyces tricolor HM10 protease and its inhibitor
The crude extract from S. tricolor HM10 culture was precipitated with ammonium sulphate, dialysed, and put onto an S200 column (89 cm × 2 cm) previously equilibrated with 20 mM Tris-HCl buffer at pH 7. The same buffer was used for the chromatography, with a flow rate of 30 mL/h. 3 mL samples were gathered. The absorbance at 280 nm as well as the protease and protease inhibitor activity at pH 9 and 7 respectively were determined for each fraction. The elution profile and the result of the partial purification are shown in Fig.3 and table 2.
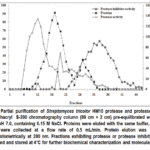 |
Figure 3: Partial purification of Streptomyces tricolor HM10 protease and protease inhibitor using Sephacryl S-200 chromatography column (89 cm × 2 cm) pre-equilibrated with 20 mM Tris-HCl, pH 7.0, containing 0.15 M NaCl. |
Table 2: Summary of partial purification of Streptomyces tricolor HM10 protease and protease inhibitor.
| Purification Step | Total Protein (mg) | Protease Activity (U) | Specific Protease Activity (U/mg protein) | Protease Recovery (%) | Protease Purification Fold | Inhibitory Activity (IU) | Specific Inhibitory Activity (IU/mg protein) | Inhibitor Recovery (%) | Inhibitor Purification Fold |
| Crude extract | 235.00 | 346 | 1.47 | 100.00 | 1.00 | 196 | 0.84 | 100.00 | 1.00 |
| Ammonium sulfate fraction | 18.87 | 273 | 15.63 | 78.90 | 10.63 | 135 | 7.15 | 68.87 | 8.51 |
| Dialysis | 14.20 | 269 | 18.94 | 77.74 | 12.88 | 128 | 9.01 | 65.31 | 10.72 |
| Sephacryl S-200 fraction | 9.37 | 183 | 19.59 | 52.89 | 13.32 | 102 | 10.88 | 52.04 | 12.95 |
Values represent the purification parameters obtained during sequential partial purification of extracellular protease and its corresponding protease inhibitor.
Gel filtration of Streptomyces tricolorHM10 crude extract on a Sephacryl S-200 column effectively resolved the sample into three distinct components (Figure 3). Fractions 1-5 showed no detectable protein or enzymatic activity. Protease activity eluted in fractions 9-12, peaking at fraction 10, whereas the majority of protein eluted in fractions 14-18 with minimal associated activity, suggesting non-active proteins. Protease inhibitor activity appeared later, in fractions 30-35, indicating a smaller molecular size relative to the protease. These results indicate that gel filtration efficiently separated the protease from its inhibitor, confirming that they are distinct molecular entities.
As shown in the table 2, we conclude that the partial purification results demonstrated the successful enrichment of both the protease and its inhibitor produced by Streptomyces tricolor HM10 through sequential ammonium sulfate precipitation, dialysis, and Sephacryl S-200 gel filtration. For the protease, the specific activity increased progressively from 1.47 U/mg in the crude extract to 19.59 U/mg after the final purification step, corresponding to a 13.32-fold purification with a final recovery of 52.89%. Similarly, the inhibitor specific activity increased from 0.84 to 10.88 IU/mg, achieving a 12.95-fold purification with 52.04% recovery. Although both total protease and inhibitory activities decreased gradually during purification due to unavoidable losses, the marked increase in specific activity and purification fold confirms the effectiveness of the purification protocol and successful removal of contaminating proteins.
Effect of pH and temperature on the protease and its inhibitor activity and stability
At 40 °C, the maximum proteolytic activity was observed and calculated as 100% relative activity. After 30 minutes of protease pre-incubation at various pH levels, a bell-shaped curve was produced, as seen in Fig. 4A. Over a wide pH range (6.0-11), protease activity was detected, peaking at pH 9.0. Between pH 6.0 and 9.0, enzyme activity climbed consistently before progressively declining.At pH 8.0, 10, 11, and 12, the relative protease activity was 93.54%, 89.24%, 18.27%, and 10.75%, respectively. Proteases that are stable and highly active in alkaline (basic) pH are well-suited for environments like detergents or certain industrial processes, where conditions are not acidic. The fact that activity peaks in this range suggests the enzyme’s structure and especially its active site is optimized to function under basic conditions.On the other hand, the low activity at acidic pH likely means the enzyme’s structure is being altered (denatured or partially disrupted) in those conditions. Changes in pH can affect the ionization of amino acids at the active site, which is critical for catalysis. If those residues lose the correct charge state, the enzyme can’t function properly.
The effect of pH on the Streptomyces tricolor HM10 protease inhibitor was also investigated. As shown in Fig.4A, maximum inhibitory activity was observed at pH 7.0. The relative activities at pH 6.0 and 8.0 were about 65 to 75% and a significant decline in activity was detected under acidic conditions, with only 28% activity retained at pH 4.0. This reduction suggests denaturation of the protease inhibitor at lower pH levels, particularly under strongly acidic conditions (pH 2.0-3.0).
pH stability studies further demonstrated that both the enzyme and its inhibitor retained substantial activity over a broad pH range. The protease remained stable at pH 8-10 for 24 h, retaining nearly 85% activity at pH 7 and 38% at pH 11, whereas the inhibitor maintained over 65% activity at pH 11 (Fig.4B).
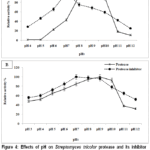 |
Figure 4: Effects of pH on Streptomyces tricolor protease and its inhibitor activity and stability. |
Optimum pH of activities. (B) pH stability of activities.
The graph shows the relative activity versus different pH ranging from 3.0 to 12 for partial purified protease and protease inhibitor.Data are expressed as mean ± SD (n = 3).
The enzyme retained 80% and 75% of its activity at 40°C and 60°C, respectively, with maximum activity observed at 50°C (Fig.5A). Regarding thermal stability, the alkaline protease remained completely stable at 50°C for 30 minutes, while only a slight reduction in activity was observed at higher temperatures, decreasing to 98% at 60°C and 90% at 70°C (Fig.5B).The protease activity was decreased with a further increase in temperature beyond 50°C due to the thermal denaturation of the 3D structure of the protein at a higher temperature.
The optimal temperature for protease inhibitor activity was determined to be 60°C (Figure 5A). Within the 40-60°C temperature range, protease activity increased. The protease inhibitor maintained 70% of its maximum activity at 80°C, however at temperatures over 60°C, a decrease in inhibitor activity was reported.
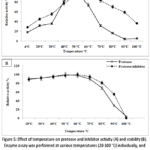 |
Figure 5: Effect of temperature on protease and inhibitor activity (A) and stability (B). Enzyme assay was performed at various temperatures (20-100 °C) individually, and relative activity was measured. |
The graph shows the relative activity at different temperatures from 20°C to 100°C for determining thermal activity and stability of protease and protease inhibitor. Data are expressed as mean ± SD (n = 3).
Storage stability
The Streptomycestricolor HM10 protease retained 80% of its activity, while its inhibitor maintained full activity (100%) during the first 21 days of storage at 4°C. Remarkably, even after 70 days, the protease and inhibitor activities remained relatively stable at approximately 60% and 90%, respectively. Furthermore, protein concentration showed minimal variation throughout the storage period (Fig. 6).
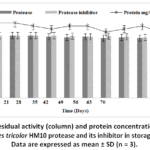 |
Figure 6: Residual activity (column) and protein concentration (line) of Streptomycestricolor HM10 protease and its inhibitor in storage condition. Data are expressed as mean ± SD (n = 3). |
Effect of metal ions and some chemicals on Streptomyces tricolorHM10 protease and its inhibitor activity
The effect of various monovalent, divalent, and trivalent metal ions on protease and its inhibitor activity is presented in Table 3. Both the protease and its inhibitor were strongly activated by Mn²⁺ and Co²⁺ at 5 mM, reaching 170% and 143% for the protease and 162% and 177% for the inhibitor, respectively. Moderate activation was observed with K⁺, Na⁺, and Fe²⁺, showing 126%, 115%, and 112% activity for the protease and 110%, 120%, and 130% for the inhibitor at the same concentration. In contrast, Hg²⁺ strongly inhibited both the enzyme and its inhibitor at 5 mM, reducing their activity to 5% and 7%, respectively, followed by Cd²⁺, Cr³⁺, and Ni²⁺, which decreased activity to 11.4%, 23.5%, and 35.1%, respectively for the protease and for the inhibitor was 17.5%, 19.1% and 22.3%. The enzyme and inhibitor were relatively stable in the presence of Mn, Mg²⁺ and Cu²⁺.
Table 3: Effect of various metal ions (5mM) on the activity of protease and protease inhibitor fromStreptomyces tricolor M10.
| Ions (5 mM) | Protease Relative activity (%) | Inhibitor Protease Relative activity (%) |
| None | 100 | 100 |
| Ca2+ | 98 | 95 |
| Mg2+ | 97 | 98 |
| Fe2+ | 112 | 130 |
| Mn2+ | 170 | 162 |
| Cu2+ | 94 | 96 |
| Ni2+ | 35.1 | 22.3 |
| Hg2+ | 5 | 7 |
| Na+ | 115 | 120 |
| K+ | 126 | 110 |
| Zn2+ | 97 | 99 |
| Cd2+ | 11.4 | 17.5 |
| Co2+ | 143 | 177 |
| Cr3+ | 23.5 | 19.1 |
The activity of the proteases and its inhibitor were determined by incubating them in the presence of various metal ions for 30 minatoptimal conditions.
The effect of chemicals on protease and its inhibitor activity
The results presented in Table 4 indicate that the nonionic surfactants Tween 80, Tween 20, and Triton X-100 reduced protease activity to 78%, 76%, and 56%, respectively, while having no significant effect on the inhibitor, which retained 95%, 89%, and 85% of its activity. This stability suggests the potential applicability of the alkaline protease in the laundry detergent industry. In contrast, both the enzyme and its inhibitor were completely inactivated by the ionic surfactant SDS, highlighting its strong denaturing effect. Furthermore, phenylmethylsulfonyl fluoride (PMSF) at 5 mM markedly inhibited protease and inhibitor activities to 37% and 22%, respectively, indicating that the protease produced by Streptomyces tricolor HM10 belongs to the serine protease family. The metalloprotease inhibitor EDTA reduced protease activity by 45% but showed no effect on the inhibitor. Similarly, iodoacetate, an alkylating agent, inhibited protease and inhibitor activities by 35% and 57%, respectively, at 5 mM, likely due to its interaction with cysteine residues and irreversible modification of sulfhydryl (-SH) groups. In contrast, β-mercaptoethanol at 5 mM completely inhibited both the enzyme and its inhibitor, suggesting that these proteins are thiol-dependent.
Table 4: Effect of some chemicals on Streptomyces tricolor HM10 protease and its inhibitor activity
| Relative activity (%) | ||
| Chemical agent Concentration (5 mM) | Protease | protease inhibitors |
| Control Surfactants
Tween 80 Tween 20 Triton X-100 |
78 76 56 |
95 86 85 |
| SDS | 0 | 0 |
| Inhibitors
PMSF EDTA Iodoacetate β-Mercaptoethanol |
37 55 65 3 |
22 99 43 0 |
Application of S. tricolor HM10 protease as a biocontrol agent against some plant pathogenic fungi
The partially purified extracellular extract enriched in alkaline protease activity was evaluated as a biocontrol agent against several pathogenic fungi using Petri dish assays at concentrations of 25-50 μL. The results, presented in Table 5, showed that Fusarium oxysporum exhibited the largest inhibition zone (80 mm) at 50 μL of enzyme, followed by Sclerotinia sclerotiorum (65 mm), while Aspergillus niger and Trichoderma spp. showed smaller inhibition zones (38 mm and 34 mm). At 25 μL, the inhibition zones for all tested fungi were reduced compared to those observed at 50 μL. Trichoderma spp. and Botrytis cinerea were less sensitive to the enzyme compared to the other fungi tested.These findings suggest that the alkaline protease has strong potential as a biocontrol agent; however, further investigations are necessary to confirm its effectiveness on a wider scale.
Table 5: Antifungal activity of protease as a biocontrol agent against some plant pathogenic fungi.
| Pathogenic fungi (mm) | Protease volume (μl) | Inhibition zone |
| Aspergillus niger | 25 50 |
24 38 |
| Fusarium oxysporum | 25 50 |
67 80 |
| Sclerotinia sclerotiorum | 25 50 |
52 65 |
| Botrytis cinerea | 25 50 |
7 28 |
| Trichoderma spp. | 25 50 |
12 34 |
Inhibitory Effect of S. tricolor HM10 protease inhibitor on Human Trypsin Activity
The S. tricolor HM10 protease inhibitor activity against Human trypsin was evaluated using a gelatin agar plate assay stained with bromophenol blue (Fig. 7). In this system, proteolytic activity is indicated by the formation of clear zones resulting from gelatin hydrolysis, whereas intact protein retains the blue coloration. Human Trypsin controls and Trypsin+inhibitor mixtures (E+I) were assessed at two inhibitor concentrations (50 and 150 mg/L) across a range of enzyme activities (5, 10, 15, and 20 U). The control wells exhibited a progressive increase in the diameter of hydrolysis zones with increasing enzyme concentration, confirming expected trypsin activity. In the presence of the inhibitor, a reduction in zone diameter was observed at all tested conditions, indicating inhibition of proteolytic activity. This effect was more pronounced at 150 mg/L compared to 50 mg/L, demonstrating a concentration-dependent inhibitory response. Furthermore, inhibition was most evident at lower enzyme units and decreased with increasing enzyme concentration, suggesting partial overcoming of inhibition at higher enzyme levels. These results indicate that the tested compound exhibits measurable and dose-dependent inhibitory activity against trypsin.
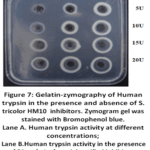 |
Figure 7: Gelatin-zymography of Human trypsin in the presence and absence of S. tricolor HM10 inhibitors. Zymogram gel was stained with Bromophenol blue. |
The inhibitory effect was found to be concentration-dependent, with higher inhibition observed at 150 mg/L compared to 50 mg/L. This suggests that the inhibitor interacts with trypsin in a dose-dependent manner, likely through the formation of enzyme-inhibitor complexes that reduce the availability of active enzyme. In addition, the results indicated that inhibition was more pronounced at lower enzyme concentrations and decreased as trypsin activity increased. This phenomenon may be explained by the stoichiometric nature of many protease inhibitors, where a fixed amount of inhibitor becomes insufficient to fully neutralize increasing amounts of enzyme. At higher enzyme concentrations, excess trypsin may remain unbound and retain catalytic activity, resulting in larger hydrolysis zones.
Amplification of S. tricolor HM10 protease and protease inhibitor genes
An initial amplification on genomic DNA was necessary to characterise the protease and protease inhibitor genes of S. tricolour HM10. As stated in the material and method chapter, we extracted genomic data in this initial stage. Spectrophotometry was used to assess the concentration of genomic DNA at 260 nm, at 250 ng/µL. The ratio of readings at 260 nm and 280 nm allowed us to estimate the amount of DNA in our preparation, which was considered satisfactory as the ratio of A260/A280 values was 1.8. The designed genes were amplified using a variety of primer combinations.
Agarose gel electrophoresis (1%) of PCR products from S. tricolor HM10 supported amplification of genes related to proteolytic functions (Fig. 8). Clear bands were observed for the serine protease gene (Lane 1), subtilase-type protease inhibitor (Lane 2), and CPBP family intramembrane metalloprotease (Lane 4), while no band was detected for the protease inhibitor I42 family protein gene (Lane 3), indicating a negative result under the present PCR conditions. The absence of amplification may be due to gene absence or sequence variation in primer-binding regions, which is commonly reported in Streptomyces species because of their genetic diversity. The detected serine protease gene agrees with previous studies showing that Streptomyces genomes frequently contain serine proteases involved in protein degradation and environmental adaptation. Subtilisin-type proteases and their inhibitors are also widely reported in actinomycetes and are known to play a role in regulating proteolytic activity. The CPBP family metalloprotease further supports the presence of membrane-associated proteolytic systems in this strain. Overall, these results support the biochemical data showing active protease production and regulation in S. tricolor HM10. These observations collectively support the role of S. tricolor HM10 as a potential source of bioactive proteases and regulatory inhibitors with applications in biotechnology and biological control.
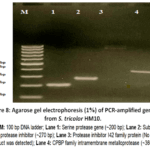 |
Figure 8: Agarose gel electrophoresis (1%) of PCR-amplified genes from S. tricolor HM10. |
Discussion
Microorganism-derived natural products constitute a rich and largely untapped reservoir of bioactive compounds and secondary metabolites with substantial therapeutic potential.20 These molecules are biosynthesized by microorganisms as part of their adaptive strategies, enabling them to survive, compete, and communicate within diverse and often hostile environments.21 Among microbial producers, Streptomyces species stand out as particularly prolific, having yielded a significant proportion of clinically relevant natural products, including antibiotics, anticancer agents, and immunosuppressants.22-26
Given the growing global demand for novel and effective therapeutics, especially in the face of drug resistance and emerging diseases, the discovery and characterization of new biomolecules remain a critical priority.27In this regard, protease inhibitors and alkaline proteases have garnered increasing attention due to their diverse and promising medical applications.28 Protease inhibitors, in particular, play a pivotal role in modulating proteolytic activity and have been successfully developed into drugs for managing chronic and life-threatening conditions such as blood clotting disorders, various forms of cancer, and viral infections.29
Proteolytic enzymes (proteases) are indispensable to cellular metabolism, participating in numerous biochemical, physiological, and regulatory processes, including protein turnover, signal transduction, and cell cycle control.30 However, dysregulation of protease activity has been implicated in a wide range of pathological conditions, including inflammation, tumor progression, and viral replication.31 Consequently, controlling protease activity through specific inhibitors represents a powerful therapeutic strategy.
Protease inhibitors exert their effects by binding to proteases and blocking their catalytic activity, thereby preventing excessive or inappropriate protein degradation.32 Clinically, they have been employed in the treatment of coagulation disorders by regulating enzymes involved in blood clot formation, in oncology by inhibiting proteases that facilitate tumor invasion and metastasis, and in antiviral therapy by targeting viral proteases essential for viral maturation and replication.33 Furthermore, proteases are often used in conjunction with non-steroidal anti-inflammatory drugs (NSAIDs) to enhance therapeutic outcomes, particularly in inflammatory conditions.28,34
Overall, the continued exploration of microorganism-derived proteases and their inhibitors holds great promise for the development of innovative and effective treatments across a broad spectrum of diseases. In line with this objective, the present study aimed to expand the existing database of actinomycete-derived proteases and their inhibitors for potential industrial applications by isolating and characterizing a novel alkaline extracellular protease and its corresponding inhibitor from Streptomyces tricolor HM10 (native to Qassim, Saudi Arabia).The possibility of using Streptomyces for protease production has been investigated because of their capacity to secrete the proteins into extra cellular media, which is generally regarded as safe with food and drug administration (GRAS). Streptomyces sps that produce proteases include S. clavuligerus, S. griseus, S. rimouses, S. thermoviolaceus, S. thermovulgaris.13-15
The present study demonstrates that ISP-4 medium is optimal for protease inhibitor production by Streptomyces tricolor HM10, indicating that nutrient composition strongly influences secondary metabolite synthesis. The early peak in inhibitor production (day 2) suggests an association with the transition to the stationary phase. This observation is consistent with previous studies showing that enzyme and metabolite production in actinomycetes is strongly dependent on incubation time and growth phase. Mehta et al.35reported that alkaline protease production by alkaliphilic actinomycetes reached a maximum after an optimal incubation period and declined thereafter. Likewise, Wahid et al.36demonstrated that extracellular protease production by Streptomyces bikiniensis is influenced by incubation time, with peak activity followed by a decrease due to changes in physiological conditions. Similarly, Gibb and Strohl37 reported that maximum protease inhibitor production in Streptomyces peucetius occurs after approximately 100 h of cultivation during the stationary phase, supporting the growth-linked regulation of enzyme-related metabolites observed in this study.
In contrast, protease production reached its maximum at day 9, particularly in the presence of casein, indicating inducible enzyme synthesis. Casein likely acts as both a nitrogen source and an inducer, enhancing protease production. This observation agrees with earlier findings reported for Streptomyces bikiniensis, Streptomyces pseudogrisiolus NRC 15, and other alkaliphilic actinomycetes,35-37 confirming that protease synthesis in Streptomyces species is often substrate-induced.
The partial purification results demonstrated the successful enrichment and separation of both the protease and its inhibitor produced by Streptomyces tricolor HM10 through sequential ammonium sulfate precipitation, dialysis, and Sephacryl S-200 gel filtration chromatography. For the protease, the specific activity increased progressively from 1.47 U/mg in the crude extract to 19.59 U/mg after the final purification step, corresponding to a 13.32-fold purification with a recovery yield of 52.89%. Similarly, the inhibitor specific activity increased from 0.84 to 10.88 IU/mg, resulting in a 12.95-fold purification with 52.04% recovery. Although the total activities decreased gradually during purification due to unavoidable losses during processing and chromatography, the substantial increase in specific activity indicates efficient removal of contaminating proteins and successful enrichment of the target biomolecules. Furthermore, the successful separation of the protease and its inhibitor by Sephacryl S-200 gel filtration chromatography confirms that they are distinct molecular entities with different molecular sizes. The earlier elution of the protease compared to the inhibitor suggests a larger molecular weight, which is consistent with previously reported molecular weights of Streptomyces proteases (30-45 kDa) and protease inhibitors (10–12 kDa).38-40 These findings demonstrate the effectiveness of size-exclusion chromatography in resolving enzyme-inhibitor systems and support the suitability of the partially purified fractions for further biochemical characterization and potential biotechnological or pharmaceutical applications.
The protease exhibited optimal activity at pH 9.0, confirming its alkaline nature. Similar pH optima have been reported for proteases for several Streptomyces strains, with optimal pH values generally ranging between 8.0 and 10 such as from Streptomyces sp. DP2, Streptomyces spp. G157, and Streptomyces sp. CN902.41-43Comparable findings were also reported for Streptomyces griseus (pronase),29,44Streptomyces sp. GS-1(optimum pH 8.5)45, Streptomyces sp. M30 (pH 9.0)46, and Streptomyces mutabilis (pH 10),47 which exhibit optimal activity in the alkaline range
However, proteases with neutral optima have also been described, such as those from Streptomyces sp. CNXK10048, highlighting the diversity within this genus. In contrast, Touioui-Boulkour et al.,49 reported optimal activity at higher alkaline pH values (10-12), indicating species-specific variation.
The protease inhibitor showed optimal activity at pH 7.0, which aligns with reports that most Streptomyces protease inhibitors are active at neutral to slightly alkaline pH (7.0-8.5),50 such as produced by Streptomyces lavendulae that shows maximum activity at pH 8.0.50 However, some inhibitors such as pepstatin exhibit optimal activity under acidic conditions (pH 3.0-5.0),51 reflecting variability depending on the target protease highlighting functional diversity among protease inhibitors across different biological sources.
Both protease and inhibitor exhibited broad pH stability, retaining activity across a wide pH range. The protease remained stable at pH 8-10 for 24 h, retaining nearly 85% activity at pH 7 and 38% at pH 11, whereas the inhibitor maintained over 65% activity at pH 11. This observation is in agreement with Si et al.,52 although differences compared to Xin et al.46 and Touioui-Boulkour et al.49 highlight species-specific adaptations.
Temperature activity analysis revealed that the protease has an optimum at 50°C and retains stability at elevated temperatures. Similar thermal characteristics have been reported for proteases from Streptomyces clavuligerus and other Streptomyces species, with optimal temperatures ranging between 55-70°C.51,53,54 The protease inhibitor also demonstrated high thermostability, maintaining significant activity up to 80°C, exceeding that reported for some Streptomyces inhibitors such as for Streptomyces spp. VL J2.55
To the best of our knowledge, not much study has been done on the stability of the partial purified Streptomyces proteases and inhibitor during storage. However, a few Bacillus-derived proteases that maintained 80-90% activity after two months at 4°C are similar to the S. tricolor HM10 protease storage stability.56,57
The activation of S. tricolor HM10 protease and inhibitor by Mn²⁺ and Co²⁺ suggests a role for metal ions in enzyme stabilization or catalytic enhancement. This result highlights the enzyme’s potential application in environments with heavy metals. Similar effects have been reported in Streptomyces and other bacterial systems.49,58-60In S. clavuligerus Mit-1, Ca²⁺ and Cd²⁺ increased protease activity up to 140%.52,59,61 Similarly, Touioui-Boulkour et al.49 observed that Ca²⁺, Mg²⁺, and Mn²⁺ enhanced protease activity from Streptomyces spp. AH4 to 200%, 150%, and 130%, respectively.52,58,62 In Geobacillus toebii LBT77, Ca²⁺ and Mg²⁺ stimulated protease activity, whereas the enzyme remained stable with Mn²⁺, Na⁺, K⁺, and Cu²⁺.51In contrast, strong inhibition by Hg²⁺ and Cd²⁺ indicates sensitivity of essential functional groups, likely thiol residues, to heavy metal interaction.
The enzyme and its inhibitor were completely inactivated by the ionic surfactant SDS, indicating its denaturing effect (Table 4). This observation agrees with previous studies on alkaline serine proteases from Streptomyces spp.,58Streptomyces spp. A652 and S. flavogriseus HS.63 However, Touioui-Boulkour et al.49 reported that SDS at 2% enhanced the activity of alkaline proteases from Streptomyces spp. AH4 up to 110%.
PMSF at 5 mM strongly inhibited the protease and inhibitor activity to 37% and 22%, respectively, indicating that S. tricolor HM10 belongs to the group of serine proteases. The metalloprotease inhibitor EDTA reduced protease activity by 45% but had no effect on the inhibitor. Iodoacetate, an alkylating agent, inhibited protease and inhibitor activity by 35% and 57%, respectively, at 5 mM, likely due to its interaction with cysteine residues and irreversible modification of sulfhydryl (-SH) groups. These findings are consistent with reports on proteases from Streptomyces spp., Streptomyces spp. A6, S. flavogriseus HS1, and Streptomyces spp. AH4.49, 54, 59, 63while complete inhibition by β-mercaptoethanol indicates the importance of thiol groups in maintaining enzyme structure. Comparable observations have been reported for proteases from Geobacillus toebiiLBT77.61
The observed antifungal activity highlights the potential of this partial purified protease as a biocontrol agent. Strong inhibition of Fusarium oxysporum is consistent with previous studies demonstrating that Streptomyces-derived proteases can suppress fungal pathogens.49,53,64 Differences in sensitivity among fungal species may reflect variations in cell wall composition or resistance mechanisms. These results suggest that the protease may have potential as a biocontrol agent. Furthermore, the partial purified protease’s stability of conservation at room temperature under field conditions indicated that it might also be used in agricultural settings.However, because the antifungal assays were performed using partially purified fractions, the contribution of additional bioactive metabolites cannot be excluded. Further studies using fully purified protease fractions are required to confirm whether the observed antifungal activity is specifically attributable to the protease activity.
The protease inhibitor exhibited concentration-dependent inhibition of trypsin, which is characteristic of reversible or competitive inhibition. Similar dose-dependent inhibition patterns have been reported for microbial protease inhibitors, particularly those from Streptomyces species.¹⁰ The reduced inhibitory effect at higher enzyme concentrations supports a stoichiometric interaction between enzyme and inhibitor.65
Finally, molecular analysis confirmed the presence of genes encoding serine protease, subtilase-type inhibitor, and metalloprotease, supporting the biochemical findings. The absence of amplification for the I42 inhibitor gene may be attributed to genetic variability or sequence divergence, which is commonly observed in Streptomyces species66,67where protease and inhibitor genes are commonly linked to biocontrol and environmental survival. These observations collectively support the role of S. tricolor HM10 as a potential source of bioactive proteases and regulatory inhibitors with applications in biotechnology and biological control.
Conclusion
Streptomyces tricolor HM10 produces a functionally active extracellular protease and a corresponding protease inhibitor with distinct biochemical properties. Both molecules are stable over a wide range of pH, temperature, and storage conditions, indicating strong environmental adaptability. The enzyme exhibits characteristics of an alkaline serine protease with industrial relevance, particularly due to its stability in surfactants and activity at elevated temperatures. The inhibitor shows specific and concentration-dependent inhibition of trypsin, suggesting regulatory and biotechnological potential. The observed antifungal activity highlights its possible application in biological control of phytopathogens. Molecular analysis confirmed the presence of key protease-related genes, supporting biochemical findings and indicating a genetically diverse proteolytic system in this strain. Collectively, these results suggest that S. tricolor HM10 is a valuable candidate for enzyme production and agricultural biocontrol applications.
Acknowledgement
The authors gratefully acknowledge Qassim University, represented by the Deanship of Graduate Studies and Scientific Research, on the financial support for this research.
Funding Sources
This research was funded by the Deanship of Graduate Studies and Scientific Research through the project number (QU-J-PG-2-2025-53507).
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable
Authors’ Contribution
- Rinad Aldhali Conceptualization, Methodology, Writing, Original Draft.
- Medhat Rehan: Data Collection, Analysis, Writing, Review & Editing.
- Imen Ben Abdelmalek: Visualization, Supervision, Project Administration Funding Acquisition.
References
- López-Otín C, Bond JS. Proteases: multifunctional enzymes in life and disease. J Biol Chem. 2008;283(45):30433-30437. doi:10.1074/jbc.R800035200
CrossRef - International Union of Biochemistry and Molecular Biology (IUBMB). Enzyme nomenclature database: EC classification system. Nucleic Acids Res. 2023;51(D1):D618-D623. doi:10.1093/nar/gkac1055
CrossRef - Puente XS, Sánchez LM, Overall CM, López-Otín C. Human and mouse proteases: a comparative genomic approach. Nat Rev Genet. 2003;4(7):544-558. doi:10.1038/nrg1111
CrossRef - Rawlings ND, Barrett AJ, Bateman A. MEROPS: the database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2012;40(D1):D343-D350. doi:10.1093/nar/gkr987
CrossRef - Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. Molecular and biotechnological aspects of microbial proteases. Microbiol Mol Biol Rev. 1998;62(3):597-635. doi:10.1128/MMBR.62.3.597-635.1998
CrossRef - Pham CTN. Neutrophil serine proteases: specific regulators of inflammation. Nat Rev Immunol. 2006;6(7):541-550. doi:10.1038/nri1841
CrossRef - Laskowski MJr, Kato I. Protein inhibitors of proteinases. Annu Rev Biochem. 1980;49:593-626. doi:10.1146/annurev.bi.49.070180.003113
CrossRef - De Clercq E. Antiviral drugs in current clinical use. J Clin Virol. 2004;30(2):115-133. doi:10.1016/j.jcv.2004.02.009
CrossRef - Flexner C. HIV protease inhibitors. N Engl J Med. 1998;338(18):1281-1292. doi:10.1056/NEJM199804303381807
CrossRef - Barka EA, Vatsa P, Sanchez L, et al. Taxonomy, physiology, and natural products of actinobacteria. Microbiol Mol Biol Rev. 2016;80(1):1-43. doi:10.1128/MMBR.00019-15
CrossRef - Goodfellow M, Williams ST. Ecology of actinomycetes. Annu Rev Microbiol. 1983;37:189-216. doi:10.1146/annurev.mi.37.100183.001201
CrossRef - Hutchings MI, Truman AW, Wilkinson B. Antibiotics: past, present and future. Curr Opin Microbiol. 2019;51:72-80. doi:10.1016/j.mib.2019.10.008
CrossRef - Taguchi S, Suzuki M, Kojima S, Miura K, Momose H. Streptomyces serine protease (SAM-P20): recombinant production, characterization, and interaction with endogenous protease inhibitor. J Bacteriol. 1995;177(22):6638-6643. doi:10.1128/jb.177.22.6638-6643.1995
CrossRef - Taguchi S, Kikuchi H, Kojima S, Kumagai I. High frequency of SSI-like protease inhibitors among Streptomyces. Biosci Biotechnol Biochem. 1993;57(3):522-526. doi:10.1271/bbb.57.522
CrossRef - Kalchev K, Rabadjiev Y, Ganchev D, et al. Study of proteases and protease inhibitors from Streptomyces Bulgar J Agric Sci. 2013;19(2):65-67
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1-2):248-254. doi:10.1016/0003-2697(76)90527-3
CrossRef - Tsuchida O, Yamagata Y, Ishizuka J, et al. An alkaline protease of an alkalophilic Bacillus sp. Curr Microbiol. 1986;14:7-12.org/10.1007/BF01568094
CrossRef - Cupp-Enyard C. Sigma’s non-specific protease activity assay: casein as a substrate. J Vis Exp. 2008;(19):899. doi:10.3791/899
CrossRef - Zhang X, Shuai Y, Tao H. Novel method for the quantitative analysis of protease activity: the casein plate method and its applications. ACS Omega. 2021;6(5):3675-3680. doi:10.1021/acsomega.0c05192
CrossRef - Bérdy J. Thoughts and facts about antibiotics: where we are now and where we are heading. J Antibiot (Tokyo). 2012;65(8):385-395.DOI: 10.1038/ja.2012.27
CrossRef - Xiao C, Xu C, Zhang J, et al. Soil microbial communities affect the growth and secondary metabolite accumulation in Bletilla striata (Thunb.) Rchb. f. Front Microbiol. 2022;13:916418. DOI: 10.3389/fmicb.2022.916418
CrossRef - Ahsan T, Chen J, Zhao X, Irfan M, Wu Y. Extraction and identification of bioactive compounds produced by Streptomyces strain KX852460. AMB Express. 2017;7:54org/10.1186/s13568-017-0351-z
CrossRef - Watve MG, Tickoo R, Jog MM, Bhole BD. How many antibiotics are produced by the genus Streptomyces?. Arch Microbiol. 2001;176(5):386-390. doi:10.1007/s002030100345
CrossRef - Takahashi Y, Nakashima T. Actinomycetes, an Inexhaustible Source of Naturally Occurring Antibiotics. Antibiotics (Basel). 2018;7(2):45. Published 2018 May 24. doi:10.3390/antibiotics7020045
CrossRef - Shih HD, Liu YC, Hsu FL, et al. Fungichromin: a substance from Streptomyces padanus. J Agric Food Chem. 2003;51:95-99. doi:10.1021/jf025879b
CrossRef - Sadeghi A, Karimi E, Dahaji PA, Javid MG, Dalvand Y, Askari H. Plant growth promoting activity of Streptomyces. World J Microbiol Biotechnol. 2012;28:1503-1509.org/10.1007/s11274-022-03380-8
CrossRef - Newman DJ, Cragg GM. Natural products as sources of new drugs. J Nat Prod. 2020;83(3):770-803.org/10.1021/acs.jnatprod.9b01285
CrossRef - Song P, Zhang X, Wang S, et al. Microbial proteases and their applications. Front Microbiol. 2023;14:1236368. doi:10.3389/fmicb.2023.1236368
CrossRef - Drag M, Salvesen GS. Emerging principles in protease-based drug discovery. Nat Rev Drug Discov. 2010;9(9):690-701. doi:10.1038/nrd3053
CrossRef - Turk B. Targeting proteases: successes, failures and future prospects. Nat Rev Drug Discov. 2006;5(9):785-799.org/10.1038/nrd2092
CrossRef - Kaur R, Rawat SS, Keshri AK, et al. An integrated approach of proteomic, cheminformatic, and in vitro drug screening. ACS Omega. 2025;10(50):61231-61246. doi:10.1021/acsomega.4c07758
CrossRef - Rawlings ND, Morton FR, Kok CY, Kong J, Barrett AJ. MEROPS: the peptidase database. Nucleic Acids Res. 2008;36(Database issue):D320-D325. doi:10.1093/nar/gkm954
CrossRef - De Clercq E. The history of antiretrovirals: key discoveries over the past 25 years. Rev Med Virol. 2009;19(5):287-299.org/10.1002/rmv.624
CrossRef - Aehle W. Enzymes in Industry: Production and Applications. 3rd ed. Weinheim, Germany: Wiley-VCH; 2007:314-320
CrossRef - Mehta VJ, Thamar JT, Singh SP. Production of alkaline protease from an alkaliphilic actinomycetes. Bioresour Technol. 2006;97:1650-1654. doi:10.1016/j.biortech.2005.07.023
CrossRef - Wahid S, Ferdous J, Anwar MN. Factors influencing production and properties of extracellular protease. Chittagong Univ J Biol Sci. 2013;4:153-163. doi:10.3329/cujbs.v4i1.13399
CrossRef - Gibb GD, Strohl WR. Physiological regulation of protease activity in Streptomyces peucetius. Can J Microbiol. 1988;34(2):187-190. doi:10.1139/m88-034
CrossRef - Sreerama L, Ramakrishna B, Sashidhar RB. Purification and properties of keratinolytic protease. J Appl Microbiol. 2001;90(3):346-352. doi:10.1046/j.1365-2672.2001.01299.x
CrossRef - Gupta R, Ramnani P. Microbial keratinases and their applications. Appl Microbiol Biotechnol. 2006;70(1):21-33. doi:10.1007/s00253-005-0255-3
CrossRef - Takahashi K, Yamada S, Nakamura K. Characterization of a novel protease inhibitor. Biochim Biophys Acta Proteins Proteom. 2005;1751(2):123-130. doi:10.1016/j.bbapap.2005.02.005
CrossRef - Bajaj BK, Sharma P. Alkali-thermotolerant extracellular protease. New Biotechnol. 2011;28(6):725-732. doi:10.1016/j.nbt.2011.05.004
CrossRef - Sampath P, Subramanian C, Chandrakasan G. Extracellular proteases from Streptomyces G157. Biotechnol Appl Biochem. 1997;26:85-90.doi.org/10.1111/j.1470-8744.1997.tb00451.x
CrossRef - Lazim H, Mankai H, Slama N, et al. Production and optimization of thermophilic alkaline protease. J Ind Microbiol Biotechnol. 2009;36(4):531-537. doi:10.1007/s10295-009-0531-3
CrossRef - Trop M, Birk Y. Specificity of proteinases from Streptomyces griseus. Biochem J. 1970;116(1):19-25. doi:10.1042/bj1160019
CrossRef - Sarkar G, Suthindhiran K. Extraction and characterization of alkaline protease. Biocatal Agric Biotechnol. 2020;25:101590. doi:10.1016/j.bcab.2020.101590
CrossRef - Xin Y, Sun Z, Chen Q. Purification and characterization of extracellular alkaline protease. J Microbiol Biotechnol. 2015;25(11):1944-1953. doi:10.4014/jmb.1507.07017
CrossRef - Mechri S, Bouacem K, Chalbi TB, et al. Taguchi design approach for alkaline thermostable protease production. J Surfactants Deterg. 2022;25(4):487-504. doi:10.1002/jsde.12567
CrossRef - Pham TV, Hua TC, Nguyen NA, Nguyen HTD. Purification and characterization of thermostable protease. Pol J Microbiol. 2024;73(2):155-165. doi:10.33073/pjm-2024-014
CrossRef - Touioui-Boulkour B, Benmrad MO, Agrebi R, et al. Two detergent-stable serine alkaline proteases. World J Microbiol Biotechnol. 2015;31:1079-1092. doi:10.1007/s11274-015-1843-6
CrossRef - Kumar S, Sharma V, Bhalla TC. Purification and characterization of protease inhibitor. Biomed Res Int. 2015;2015:963041. doi:10.1155/2015/963041
CrossRef - Umezawa H, Aoyagi T, Morishima H, et al. Pepstatin, a new pepsin inhibitor. J Biol Chem. 1970;245(15):3975-3978. doi:10.1016/S0021-9258(70)90026-4
- Si JB, Jang EJ, Charalampopoulos D, Wee YJ. Protease from Bacillus subtilis FBL-1. Biotechnol Bioprocess Eng. 2018;23:176-182. org/10.1007/s12257-017-0495-3
CrossRef - Thumar JT, Singh SP. Secretion of alkaline protease from Streptomyces clavuligerus. Braz J Microbiol. 2007;38:766-772. doi:10.1590/S1517-83822007000400033
CrossRef - Singh AK, Chhatpar HS. Purification of antifungal protease. J Basic Microbiol. 2011;51:424-432. doi:10.1002/jobm.201000310
CrossRef - Marathe K, Kasar S, Chaudhari A, Maheshwari V. Protease inhibitor from Streptomyces Process Biochem. 2016;51(10):1650-1663. doi:10.1016/j.procbio.2016.07.005
CrossRef - Kotb E, Azzaz HH, Hamed MM. Isolation of alkaline protease-producing bacteria. Fermentation. 2023;9(7):637. doi:10.3390/fermentation9070637
CrossRef - Wang K, Tian Y, Zhou N, Li Y, Zhang X. Stability and application of proteases. Process Biochem. 2018;74:10-20. doi:10.1016/j.procbio.2018.08.035
CrossRef - El-Khonezy MI, El-Sayed HS, El-Hersh MS, Abou-Elela GM. Partial purification of alkaline serine protease. World Appl Sci J. 2015;33:831-842.doi: 10.5829/idosi.wasj.2015.33.05.14588
- Yum DY, Lee SG, Kim JH. Alkaline serine protease from Biosci Biotechnol Biochem. 1994;58:470-474. doi:10.1271/bbb.58.470
CrossRef - Patel Y, Gupte A, Gupte S. Production of alkaline protease from Bacillus cereus Int J Curr Microbiol Appl Sci. 2018;7(8):587-600. doi:10.20546/ijcmas.2018.708.064
CrossRef - Thebti W, Riahi Y, Belhadj O. Thermostable protease from Geobacillus toebii.Biomed Res Int. 2016;2016:9178962. doi:10.1155/2016/9178962
CrossRef - Hou E, Xia T, Zhang Z, Mao X. Alkaline protease from Micrococcus sp. J Ocean Univ China. 2017;16:319-325. doi:10.1007/s11802-017-3207-x
CrossRef - Ghorbel S, Kammoun M, Soltana H, et al. Proteases from Streptomyces flavogriseus. Biomed Res Int. 2014;2014:345980. doi:10.1155/2014/345980
CrossRef - Al-Askar AA, Rashad YM, Hafez EE, et al. Alkaline protease for controlling Rhizoctonia root rot. Biotechnol Biotechnol Equip. 2015;29(3):457-462. doi:10.1080/13102818.2015.1015446
CrossRef - Quintero D, Bermudes D. Method for protease inhibitor detection. J Microbiol Methods. 2014;100:105-110. doi:10.1016/j.mimet.2014.02.019
CrossRef - Barka EA, Vatsa P, Sanchez L, et al. Actinobacteria and bioactive compounds. Microbiol Mol Biol Rev. 2020;84(1):e00019-19. doi:10.1128/MMBR.00019-19
CrossRef - Nguyen HT, Chau MN, Kim JC. Genomic insights into Streptomyces. Front Microbiol. 2021;12:745812. doi:10.3389/fmicb.2021.745812
Abbreviations
| ANOVA | Analysis of Variance | PDA | Potato Dextrose Agar |
| bp | Base pair | PIU | Protease Inhibitor Unit |
| BSA | Bovine Serum Albumin | PMSF | Phenylmethylsulfonyl Fluoride |
| CPBP | CAAX Protease and Bacteriocin Processing protein | rpm | Revolutions Per Minute |
| DNA | Deoxyribonucleic Acid | SDS | Sodium Dodecyl Sulfate |
| dNTPs | Deoxynucleotide Triphosphates | SD | Standard Deviation |
| EC | Enzyme Commission | SMF | Submerged Fermentation Medium |
| EDTA | Ethylenediaminetetraacetic Acid | SSI | Streptomyces Subtilisin Inhibitor |
| HM10 | Strain designation of Streptomyces tricolor | SIL | SSI-Like Inhibitors |
| I42 | Protease Inhibitor Family I42 | TAE | Tris-Acetate-EDTA Buffer |
| ISP-4 | International Streptomyces Project Medium 4 | TCA | Trichloroacetic Acid |
| kDa | Kilodalton | Tris-HCl | Tris(hydroxymethyl)aminomethane Hydrochloride |
| MgCl₂ | Magnesium Chloride | UV | Ultraviolet |
| MnCl₂ | Manganese Chloride | NaCl | Sodium Chloride |
| PCR | Polymerase Chain Reaction |

